
肺腺癌增强子调控miRNA前馈环路的识别与功能分析
2019-02-28 03:26李志学梁子涵贾承霖漆宇骋岳俊杰郭志云
生物技术通讯 2019年6期
李志学,梁子涵,贾承霖,漆宇骋,岳俊杰,郭志云
1.西南交通大学 生命科学与工程学院,四川 成都610031;2.军事科学院 军事医学研究院 生物工程研究所,北京100071
肺癌在世界范围的发病率和致死率都高居各类癌症之首,分别达11.6%和18.4%[1]。肺癌分为小细胞癌和非小细胞癌,非小细胞癌又分为鳞状细胞癌、腺细胞癌等亚型,其中肺腺癌最为常见。肺腺癌治疗方法主要包括手术、放疗、化疗和靶向治疗,但仅靠这些疗法还无法保证彻底治愈[2]。因此,关于肺腺癌发生发展分子机制及调控机理的研究,仍是学术界关注的热点。
在人类进化演变的过程中,形成了纷繁复杂又高度有序的基因网络调控结构,它们由一定的模序(motif)元件所构成,在生物体内发挥着重要的基因调节作用[3]。其中,前馈环路(feed-for⁃ward loops,FFLs)是一种重要的循环线性模序,它由2个输入调节因子P1和P2,以及1个输出因子P3 构成。P1 可以调节P2,也可以与P2 共同调节P3,即P3 所受到的网络调节的影响是P1、P2 元件综合作用的结果。
增强子是真核细胞中普遍存在的顺式调控元件,可以通过招募转录因子(transcription fac⁃tor,TF),进而结合多种激活因子,形成多蛋白复合物,使染色质成环,远端调控启动子。通过对增强子染色质的修饰和重塑等作用,促进基因的转录[4]。此外,较多证据表明,增强子与microRNA(miRNA)间同样存在复杂的调控关系。Xiao 等[5]通过删除HEK293T细胞系中的miR-24-1 增强子基因座,发现miR-24-1的表达量下降,其邻近基因不能再被激活,表明miR-24-1 邻近基因的转录激活依赖于完整增强子的存在。此外,miR-24-1可激活增强子RNA(eRNA)表达,改变组蛋白修饰,增加增强子位点p300和RNA PolⅡ的富集,从而激活转录。Suzuki 等证明增强子可以通过募集Drosha和DGCR8 这2个在miRNA初始转录本(primary miRNA,pri-miRNA)剪接过程中发挥重要作用的蛋白,调控成熟的miRNA在细胞中的水平,进而影响基因的表达[6]。目前,关于肺腺癌前馈环路的研究,多集中于miRNA-TF-基因所形成的调控网络,而增强子在该调控网络中发挥什么样的作用、如何发挥作用,还不够清晰。
考虑到前馈环路模型的多样性和灵活性,我们在此将其引入肺腺癌分子机制的研究,通过数据筛选和处理,期望进一步了解在肺癌腺产生过程中,增强子、转录因子、miRNA、基因之间存在的调控关系。综合多组学数据,本研究构建了TF-增强子-miRNA的前馈环路,并探讨该前馈环路的功能,为探究肺腺癌发生的分子机制及其调控机理提供新的思路,也为以网络标志物为主的肺腺癌诊断提供理论依据。
1 材料与方法
1.1 肺腺癌增强子数据的获取
增强子数据下载自HACER[7]数据库。该数据库通过GRO_seq和PRO_seq 整合FANTOM与其他转录非稳定RNA的增强子,是目前最为全面的增强子数据库。我们从HACER 数据库下载肺腺癌A549细胞增强子3870个。
1.2 TF-增强子作用关系的识别
从Cistrome DB[8]和Unibind[9]数据库下载获取A549细胞系中的97个转录因子的Chip-Seq 数据集(.bed 文件),作为转录因子与DNA的结合位点信息。当转录因子的结合位点包含于同一染色体的增强子区域时,认为该转录因子对该增强子具有调控关系。
1.3 增强子-miRNA作用关系的识别
miRNA位置信息下载自miRBase 数据库[10],miRNA转录起始位点数据下载自FANTOM 数据库,人类蛋白质编码基因注释数据下载自GEN⁃CODE 数据库。根据先前的增强子调控miRNA研究[11],对每个增强子的上游或下游,分别找到距增强子中心最近的miRNA转录起始位点和同侧的所有蛋白质编码基因转录起始位点。将增强子中心与miRNA转录起始位点间的距离定为M,增强子中心与同方向蛋白质编码基因转录起始位点间的距离定为G。以调控评分公式计算score值,设定score值为0~0.2的增强子与miRNA之间存在调控关系。
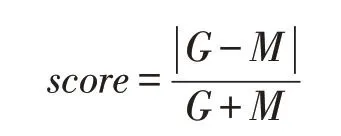
1.4 TF-miRNA作用关系的识别
利用上述获取的转录因子、miRNA数据,如转录因子的结合位点包含于miRNA转录起始位点上游10 kb 及下游1 kb,认为该转录因子对该miRNA具有调控作用[12]。
1.5 TF-增强子-miRNA前馈环路的合并与多重假设检验
将上述3 类调控关系进行组合筛选,采用超几何检验获取非随机性的前馈环路。公式如下:

其中,M为miRNA总数,N为某一转录因子所调控的miRNA总数,k为某一增强子所调控的miRNA总数,x为该转录因子所调控的miRNA与该增强子所调控的miRNA的交集。依据Benjamini-Ho⁃chberge[13]方法,利用FDR值对该超几何检验的概率值进行修正,并计算q值,将q≤0.05的前馈环路作为有生物学意义的前馈环路。
1.6 在前馈环路中提取关键性miRNA
miRNA都能够对基因表达起调控作用,从构建的TF-增强子-miRNA前馈环路中选择参与多个前馈环路的miRNA,获取参与前馈环路的每个miRNA的表达量,将miRNA高于平均miRNA表达的作为前馈环路关键性的miRNA。前馈环路关键性miRNA表达量下载自FANTOM5 数据库[14]。
1.7 前馈环路关键性miRNA的靶基因识别与富集分析
从mirTarBase[15]数据库下载经实验证实的miRNA靶基因,采用R软件包clusterProfiler 对miRNA靶基因进行Gene Ontology(GO)与KEGG Pathway 功能富集分析。
1.8 前馈环路关键性miRNA的靶基因生存分析
利用KMplot[16]生存分析在线工具,选择TF-增强子-miRNA前馈环路关键靶基因进行临床样本的生存分析。
2 结果与讨论
2.1 转录因子、增强子、miRNA作用关系筛选
通过上述数据库搜索,获取了97个转录因子的DNA 结合位点信息、3870个肺腺癌组织特异性活性增强子区域信息、4801个miRNA的转录起始位点信息和80 455个蛋白质编码基因的转录起始位点信息。最终,我们识别了227个TF-增强子作用关系,涉及61个转录因子和66个增强子;6973个增强子-miRNA作用关系,涉及3767个 增强子和1262个miRNA;10 715个TF-miRNA作用关系,涉及89个转录因子和1367个miRNA。
2.2 肺腺癌前馈环路构建
通过对上述作用关系进行整合,最终共获得36个肺腺癌TF-增强子-miRNA前馈环路(表1),其中增强子(chr11:355877~356724)参与的前馈环路数目最多,为7个。本研究构建的前馈环路涉及增强子,增强子对miRNA的调控全为正调控,由此构建的前馈环路均为连贯前馈环路[3]。
在这些前馈环路中,平均每个miRNA涉及2.769个转录因子,其中大于等于3个转录因子的miRNA有hsa-miR-6734、hsa-miR-4677、hsa-miR-6743、hsa-let-7i、hsa-miR-210、hsa-miR-4492、hsa-miR-6760。hsa-miR-210 已被证实与肺癌相关[17]。这些miRNA在肺癌发生发展的过程中可能起关键作用,例如调控肺癌相关基因的表达,提高肺癌细胞的转移性、浸润性或异质性等。
2.3 肺腺癌前馈环路关键miRNA的GO与KEGG功能富集分析
从mirTarBase 数据库获取了36个TF-增强子-miRNA前馈环路中miRNA的所有靶基因,共计2404个,利用R 软件包clusterProfiler 对所有靶基因进行了GO 功能富集。我们发现肺腺癌前馈环路在细胞周期正调控、细胞分化、蛋白质稳定性调控、能量代谢和糖代谢等方面具有重要功能(图1),这与文献报道的癌细胞基因功能一致。通过功能富集分析,我们发现了在肺癌细胞中起关键作用的多效基因TP53、IGF1、TPR、CREB、CASP3、STAT5B、APP、USP19 等,其中TP53、IGF1、TPR、CREB、CASP3、STAT5B、APP、USP19 已被证实与肿瘤密切相关,如TP53[18]和CREB基因已被证实可显著影响肺腺癌的增殖。Illiano 等采用脂联素干预A549 肺腺癌细胞,发现脂联素可抑制CREB基因表达,显著降低肺腺癌细胞的增殖[19]。

图1 肺腺癌全前馈环路靶基因GO 功能富集结果
对靶基因进行KEGG 通路富集分析(图2),发现与肺腺癌前馈环路miRNA靶基因广泛参与糖代谢、甘氨酸/丝氨酸/苏氨酸代谢、丙氨酸/天冬氨酸/谷氨酸代谢、半胱氨酸/甲硫氨酸代谢等通路。该结果说明肺腺癌前馈环路在大分子化合物代谢通路中起关键作用,该结果与癌细胞生理特征一致,其中涉及的关键基因有PGAM1、PGAM4、LDH4、LDHB、AGXT2 等。
为探究上述肺腺癌TF-增强子-miRNA前馈环路关键靶基因对肺腺癌患者的影响,我们对其中参与富集功能(≥2个)的多效基因进行生存分析。选择具有表达活性差异的基因发现,TP53和CREB1在高表达时会使肺腺癌患者的生存率降低,而STAT5B和TPR在低表达时会使肺腺癌患者的生存率降低(图3)。因此与多效基因TP53、CREB1、STAT5B、TPR 有关的前馈环路可能对肺腺癌的发展、转移有重要影响。
2.4 肺腺癌前馈环路关键miRNA表达特征
依据FAMTOM5 提供的miRNA表达量数据,发现上述6个miRNA中,hsa-let-7i-3p、hsa-miR-210-5p、hsa-miR-6734-5p与hsa-miR-4677-5p在体内的表达量显著高于前馈环路涉及到的13个miRNA表达量的平均值,暗示这4个miRNA可能是肺腺癌的关键性miRNA。其中hsa-miR-6734参与了7个前馈环路,显著高于表达量平均值2.769,共涉及7个重要的转录因子ETS1、GABPA、PBX3、TEAD4、YY1、ZBTB33、DNase。其中的YY1已有文献报道与肿瘤发生有关[20]。

表1 36个肺腺癌TF-增强子-miRNA前馈环路

图2 肺腺癌全前馈环路靶基因KEGG 通路富集结果
2.5 4个关键miRNA靶基因功能分析

图3 多效基因TP53(A)、CREB1(B)、
分别对上述4个关键miRNA靶基因进行GO功能分析和KEGG 通路分析。一方面我们获取了这4个关键miRNA的功 能活性,hsa-miR-6734 具有强的核酸转录抑制活性,hsa-miR-210 具有强的分解代谢活性,hsa-let-7i 具有强的丝氨酸/苏氨酸激酶活性,而hsa-miR-4677 无明确的富集功能。另一方面,构建4个关键miRNA的基因调控网络,发现hsa-miR-210 调控细胞分解代谢和应激反应功能的关键靶基因PLK1、APC、BNIP3、ATG7、SIN3A,hsa-miR-6734 调 控 各 类 复 合 物(NuRD、CHD 类等)功能的关键靶基因MTA1、CSNK2A1、RBBP4、ZBTB7A,hsa-let-7i细胞周期正调节功能的关键靶基因IGF1、PDGFB、CDKN1A、CCND1、EDN1和CCNT2。
2.6 结论
我们探讨了TF-增强子、增强子-miRNA、TFmiRNA作用关系,并识别了肺腺癌细胞系相关的36 条TF-增强子-miRNA前馈环路,涉及22个转录因子、13个增强子、13个miRNA。在功能富集和通路富集方面,我们发现肺腺癌细胞TF-增强子-miRNA前馈环路靶基因显著富集于与癌症密切相关的生物功能上,如细胞周期正调控、细胞分化、蛋白质稳定性调控、能量代谢和糖代谢等,其中涉及的关键基因已被很多研究证实在癌症细胞中特异表达。KEGG 通路富集结果显示前馈环路靶基因富集于肺癌代谢相关信号通路。结合miRNA的参与环路数和表达量,我们筛选了前馈环路关键miRNAhsa-miR-6734、hsa-miR-210、hsa-miR-4677和hsa-let-7i,探讨了它们与基因的调控关系网络。另外,我们对参与富集功能多的多效基因进行肺腺癌生存分析,发现前馈环路的多效靶基因TP53、CREB1、STAT5B、TPR的表达差异对患者肺癌的生存发展有重要影响。本研究揭示了肺腺癌相关前馈环路的特点,为以后肺腺癌前馈环路研究提供理论与方法参考。
猜你喜欢
上海金属(2021年6期)2021-12-02
河北工业大学学报(2021年4期)2021-09-23
昆明医科大学学报(2021年3期)2021-07-22
生物学通报(2019年3期)2019-02-17
电脑知识与技术(2018年19期)2018-11-01
中成药(2018年7期)2018-08-04
现代防御技术(2017年2期)2017-05-13
西南军医(2016年3期)2016-01-23
中国当代医药(2015年17期)2015-03-01
中华皮肤科杂志(2014年3期)2014-12-19
