
姜黄素联合KLF8基因siRNA调控JAK2/STAT3信号通路对乳腺癌细胞生长抑制作用的研究
2019-02-22 00:44李庆辉张成娟
安徽医科大学学报 2019年1期
李 新 ,牛 冰 ,李庆辉 ,张成娟
Krüppel样转录因子8(KLF8)是Krüppel样转录因子(Krüppel like transcription factor,KLFs)家族成员之一,是表达广泛的一个Krüppel样转录因子,参与细胞的增殖、分化、凋亡、迁移、细胞外基质形成等多种生物学过程[1],研究[2-3]显示,包括乳腺癌在内的多种肿瘤中KLF8呈现出过表达,而抑制KLF8后可降低肿瘤细胞生长,提示抑制乳腺癌中KLF8的表达可能影响其发生发展,但目前研究尚不清楚。
姜黄素(Curcumin)是姜黄的根茎中提取出来的一种酚性色素,具有明显的抗肿瘤作用,有研究[4]显示,姜黄素可明显抑制多种肿瘤的生长及诱导凋亡。抑制KLF8基因表达与姜黄素联合用于乳腺癌是否产生协同治疗作用尚不清楚。因此,该研究通过RNA干扰抑制KLF8基因表达及姜黄素单独或共同处理乳腺癌细胞,检测对癌细胞活力和凋亡的影响,并进一步研究对其生物学特性影响的可能分子机制。
1 材料与方法
1.1主要试剂和仪器小牛血清及RPMI1640培养基购自美国Gibco公司;姜黄素购自美国Sigma公司;AG490购自美国Biosource公司;细胞计数试剂盒(cell counting kit-8,CCK-8)试剂盒和二喹啉甲酸(BCA)试剂盒购自上海碧云天公司;LipofectamineTM2000转染试剂盒、Annexin V-FITC/PI细胞凋亡试剂盒购自美国Invitrogen公司;KLF8、磷酸化的蛋白酪氨酸激酶2(phosphorylated Janus kinase2,p-JAK2)、磷酸化的信号转导与转录因子3(phosphorylated signal transducers and activators of transcription 3,p-STAT3)、细胞周期素D1(Cyclin D1)和B细胞淋巴瘤/白血病-2(B cell lymphoma/lewkmia-2,Bcl-2)抗体购自美国Abcam公司;酶标仪及流式细胞仪均购自美国Bio-Rad公司。
1.2细胞及其培养人乳腺癌MCF-7细胞购自中国科学院上海细胞库;MCF-7细胞为贴壁细胞,细胞在含有小牛血清及双抗的RPMI1640培养基中, 5% CO2、37 ℃饱和湿度培养箱中传代培养。实验选择生长至对数期的细胞。
1.3细胞转染MCF-7细胞进行常规消化后收集细胞,接种细胞于6孔板,转染前1 d更换培养液(含有血清,不含抗生素),待细胞达70%~90%的生长密度时通过脂质体LipofectamineTM2000转染试剂盒将合成的干扰KLF8表达的siRNA(KLF8-siRNA)转染细胞,同时转染无干扰作用的siRNA作为阴性对照组,并设置只加入脂质体的为空白对照组,转染过程严格参照试剂盒说明,收集转染48 h的细胞用于实验研究。
1.4转染KLF8-siRNA的MCF-7细胞KLF8的表达通过Western blot检测转染KLF8-siRNA的MCF-7细胞KLF8的表达。简要步骤如下:PBS洗涤转染48 h的细胞,离心后弃掉上清,加入全细胞蛋白裂解液于冰上反应30 min,离心收集上清液,上清液即为细胞总蛋白。BCA试剂盒对蛋白进行定量,蛋白经100 ℃变性,取等量变性蛋白进行SDS-PAGE分离,电泳结束后切下含有目的蛋白的胶通过电转移至PVDF膜,5%脱脂奶粉在室温条件下封闭转好的PVDF膜2 h,封闭后置入适当比例的KLF8和GAPDH一抗溶液中,4 ℃缓慢摇动过夜,洗膜,加入二抗,室温反应2 h,ECL发光后暗室中将X光片曝光,冲洗X光片,拍照扫描,成像分析软件分析KLF8的相对表达量。
1.5CCK-8法检测姜黄素联合KLF8基因siRNA对MCF-7细胞活力的影响以每孔100 μl(约1×104个细胞)接种生长至对数期的MCF-7细胞于96孔板,用于调零组细胞的仅仅加入细胞培养液,细胞长满50%~80%孔底时参照上述方法进行转染,并随机分为阴性对照组(转染无干扰作用的siRNA)、KLF8-siRNA(转染合成的干扰KLF8表达的siRNA)、姜黄素组(40 μmol/L姜黄素处理细胞)和KLF8-siRNA+姜黄素组(在转染KLF8-siRNA的基础上加入姜黄素),每组设置5个平行孔,培养48 h后,加入10 μl CCK-8试剂至每孔中,利用调零组调零,酶标仪测定吸光度值(optical density,OD),以OD值反映细胞活力,可以间接反映出细胞的增殖能力。实验重复3次。
1.6流式细胞仪检测姜黄素联合KLF8基因siRNA对MCF-7细胞凋亡的影响分组及处理方法同1.5,PBS洗涤各组细胞,Binding缓冲液重悬细胞,利用Annexin V-FITC和PI荧光染色,室温避光反应15 min,流式细胞仪进行检测。实验重复3次。
1.7姜黄素联合KLF8基因siRNA抑制JAK2/STAT3信号通路对乳腺癌细胞活力及凋亡的影响AG490作为JAK2/STAT3信号通路抑制剂,加入浓度为20 μmol/L,通过CCK-8法及流式细胞仪检测用KLF8-siRNA+姜黄素+AG490处理细胞相对于KLF8-siRNA+姜黄素处理的细胞活力及凋亡率情况,Western blot检测p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表达,方法同1.4。

2 结果
2.1KLF8-siRNA转染MCF-7细胞后KLF8的表达KLF8-siRNA转染MCF-7细胞后KLF8的表达结果见图1。与空白对照组(0.311±0.035)比较,KLF8-siRNA组(0.102±0.010)KLF8的表达显著降低(P<0.05),而阴性对照组(0.302±0.032)和空白对照组KLF8的表达差异无统计学意义(P>0.05)。

图1 KLF8-siRNA转染MCF-7细胞后KLF8的表达
2.2姜黄素联合KLF8-siRNA对MCF-7细胞活力的影响CCK-8检测结果显示:KLF8-siRNA组(0.562±0.048)和姜黄素组(0.626±0.052)的细胞活力均显著低于阴性对照组(0.873±0.074)(P<0.05),高于KLF8-siRNA+姜黄素组(0.385±0.041)(P<0.05)。
2.3姜黄素联合KLF8-siRNA对MCF-7细胞凋亡的影响流式细胞仪检测结果显示:KLF8-siRNA组(12.12±1.02)%和姜黄素组(10.77±0.96)%的细胞凋亡率均显著高于阴性对照组(1.56±0.16)%(P<0.05),低于KLF8-siRNA+姜黄素组(18.79±1.35)%(P<0.05)。见图2。
2.4姜黄素联合KLF8-siRNA对MCF-7细胞p-JAK2、p-STAT3、CyclinD1和Bcl-2表达的影响Western blot检测结果显示:KLF8-siRNA组和姜黄素组Cyclin D1、Bcl-2、p-JAK2和p-STAT3的蛋白表达均显著低于阴性对照组(KLF8-siRNA组:P<0.05;姜黄素组:P<0.05),高于KLF8-siRNA+姜黄素组(KLF8-siRNA组:P<0.05;姜黄素组P<0.05)。四组间JAK2和STAT3的蛋白表达差异无统计学意义。见表1、图3。
2.5姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号对MCF-7细胞活力及凋亡的影响姜黄素、KLF8-siRNA及JAK2/STAT3信号抑制剂AG490共同处理细胞后,细胞OD值及凋亡率检测结果显示:与KLF8-siRNA+姜黄素组[(0.422±0.043)、(16.68±1.21)%]比较,KLF8-siRNA+姜黄素+AG490组OD值(0.301±0.032)显著降低(P<0.05),凋亡率显著升高[(21.15±1.54)%](P<0.05)。见图4。
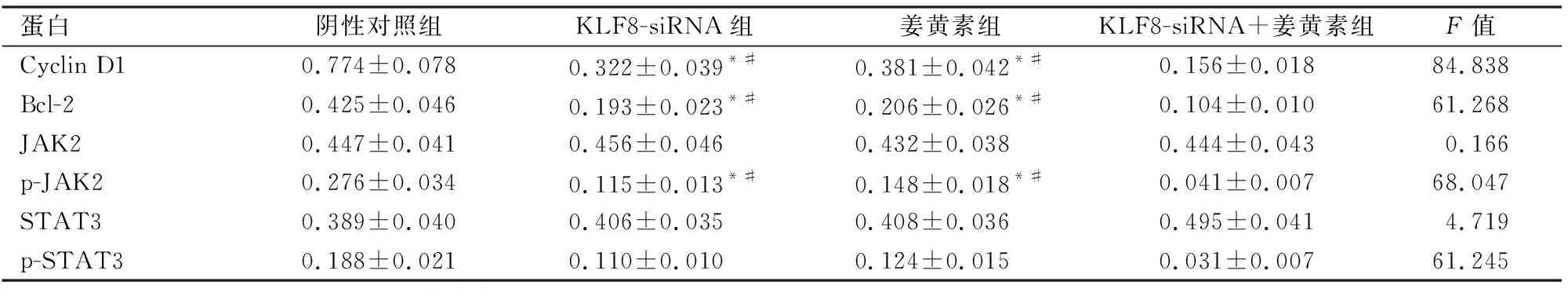
表1 姜黄素联合KLF8-siRNA对MCF-7细胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表达的影响
与阴性对照组比较:*P<0.05;与KLF8-siRNA+姜黄素组比较:#P<0.05

图2 姜黄素联合KLF8-siRNA对MCF-7细胞凋亡的影响

图3 姜黄素联合KLF8-siRNA对MCF-7细胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表达的影响
A:阴性对照组;B:KLF8-siRNA组;C:姜黄素组;D:KLF8-siRNA+姜黄素组
2.6姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号对MCF-7细胞p-JAK2、p-STAT3、CyclinD1和Bcl-2表达的影响姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号后MCF-7细胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表达结果见表2、图5。与KLF8-siRNA+姜黄素组比较,KLF8-siRNA+姜黄素+AG490组p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表达均显著降低(P<0.05)。
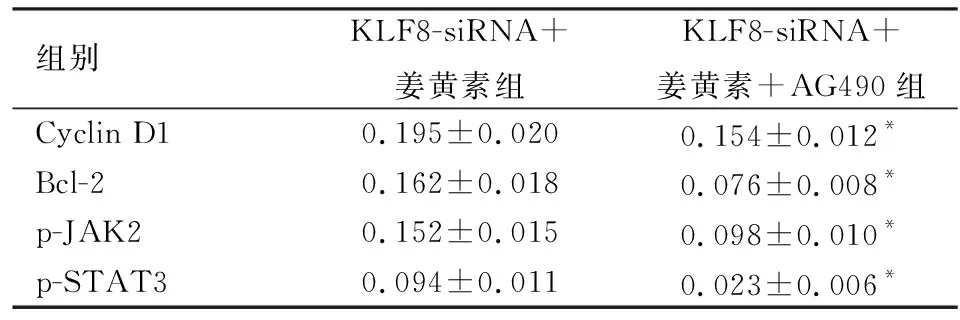
表2 姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号对MCF-7细胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表达的影响
与KLF8-siRNA+姜黄素组比较:*P<0.05
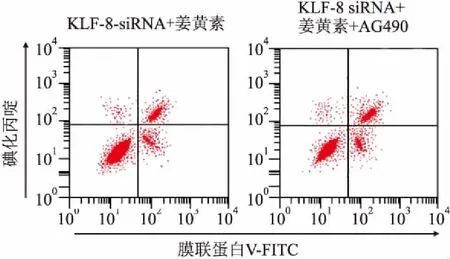
图4 姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号对MCF-7细胞活力及凋亡的影响
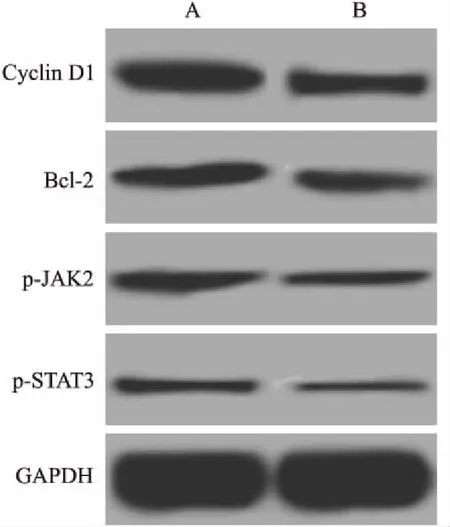
图5 姜黄素联合KLF8-siRNA抑制JAK2/STAT3信号对MCF-7细胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表达的影响
A:KLF8-siRNA+姜黄素组;B:KLF8-siRNA+姜黄素+AG490组
3 讨论
研究[5]显示,在多种肿瘤中,KLFs家族的多个成员与癌基因相关。KLF8是KLFs家族成员之一,在上皮细胞-间充质转化(epithelial cell mesenchyme transformation,EMT)、细胞致癌性分化、细胞侵袭和迁移、周期循环等过程中均有重要作用,且在多种肿瘤中过表达,其过表达与疾病不良预后及发生相关[6]。也有研究[7]指出抑制KLF8的表达可降低肿瘤的发生发展,如抑制肾癌细胞KLF8表达可降低癌细胞增殖、侵袭及诱导细胞凋亡。KLF8在乳腺癌中的研究相对较少,研究[8]显示,KLF8在乳腺癌中表达上调,可通过活化基质金属蛋白9促进乳腺癌的侵袭及转移。本研究旨在抑制KLF8对乳腺癌增殖凋亡的影响。我国中药资源丰富,中药提取物在肿瘤治疗的研究受到广泛关注,姜黄素是姜黄的根茎中提取出来的一种酚性色素,目前在肿瘤中的作用有大量研究,研究[9]显示,姜黄素可降低乳腺癌的增殖、侵袭、迁移,阻滞细胞周期和诱导细胞的凋亡,但联合使用姜黄素和KLF8的siRNA是否能更有效地治疗乳腺癌还未清楚。
鉴于有研究[2]已证实乳腺癌中KLF8存在高表达,本研究检测抑制KLF8表达对乳腺癌细胞的影响。由于RNA干扰技术能在基因的转录和翻译水平及染色质水平调节基因的表达,且表现出很强的序列特异性、有效性,在多种肿瘤中研究基因功能时被应用[10],因此本研究也选用RNA干扰技术抑制乳腺癌中KLF8的表达。将KLF8的siRNA和姜黄素共同处理乳腺癌细胞,结果显示KLF8-siRNA和姜黄素均能明显抑制乳腺癌细胞活力和诱导细胞凋亡,这提示KLF8基因和姜黄素能协同用于乳腺癌的防治。信号转导与转录因子3(signal transducers and activators of transcription 3, STAT3)是STATs家族的一员,与细胞增殖、凋亡等生物学特性密切相关,在多种肿瘤中出现异常活化,STAT3的异常活化可引起细胞异常增殖分化,且凋亡受到抑制,目前已被确认为癌基因,STAT3引起癌变的机制主要是通过激活CyclinD1、Bcl-2、Survivin等靶基因的一些产物表达实现[11]。CyclinD1是一个细胞周期核因子,可促进细胞的增殖[12]。Bcl-2是Bcl-2家族成员之一,抑制其表达可诱导细胞的凋亡[13]。JAK2处在STAT3上游,JAK2激酶抑制剂AG490可抑制JAK2/STAT3信号通路,从而使细胞增殖受到抑制,AG490可抑制包括乳腺癌在内的多种肿瘤的发生发展[14]。本研究显示,姜黄素和KLF8的siRNA均能下调p-JAK2和p-STAT3及靶基因CyclinD1和Bcl-2表达,两者联合下调更明显,抑制JAK2/STAT3信号通路后,相对于联合使用姜黄素和KLF8的siRNA对细胞的抑制作用和凋亡促进作用更明显。
综上所述,下调KLF8基因表达和姜黄素均能通过抑制JAK2/STAT3信号通路降低乳腺癌细胞活力,诱导细胞凋亡,两者联合对细胞活力及凋亡的影响作用更强。该研究可能为乳腺癌的治疗提供了新的途径,值得进一步深入探讨。
猜你喜欢
新乡医学院学报(2021年12期)2022-01-21
当代陕西(2020年9期)2020-08-04
浙江医学(2020年13期)2020-07-30
Medical Data Mining(2019年2期)2019-07-16
浙江医学(2018年16期)2018-09-08
中成药(2018年2期)2018-05-09
中成药(2018年3期)2018-05-07
中国工程咨询(2016年6期)2016-01-31
中国卫生(2015年7期)2015-11-08
西南军医(2015年1期)2015-01-22
