
航天诱变技术在小麦育种上的应用
2019-02-18 05:44张福彦张建伟程仲杰陈晓杰齐红志杨保安范家霖
核农学报 2019年2期
张福彦 张建伟 程仲杰 陈晓杰 齐红志 杨保安,∗范家霖 崔 龙
(1 河南省科学院同位素研究所有限责任公司/河南省核农学重点实验室,河南郑州 450015;2河南省农业科学院农业经济与信息研究所,河南郑州 450002)
小麦(Triticum aestivum L.)是我国重要的粮食作物之一。 优异的种质资源是小麦育种改良的基础,种质资源创新在小麦育种和提高小麦产量中发挥关键性的作用[1]。 大量研究表明,诱变育种能创造传统杂交育种无法短期得到的新的优良品种或特异优良种质材料,已成为小麦种质资源创新的重要途径之一,也是核技术农业应用研究的重要组成部分[2-3]。
诱变育种是指利用物理因素(X 射线、γ 射线、中子、电子束等具有辐射能的射线)、化学因素(甲基磺酸乙酯、硫酸二乙酯、叠氮化钠等化学诱变剂)和某些生理活性物质等对植物进行人工诱变进而产生新变异、新品种的育种技术[4-5]。 1928年,Stadler[6]首次证明X 射线对大麦具有诱发突变的效应,之后研究学者利用辐射诱变技术在农作物[7-8]、经济作物[9]、观赏性植物[10]等领域选育了多个新品种[11-12]。 我国诱变育种起步于20 世纪50年代后期,虽然起步较晚,但发展迅速。 据联合国粮农组织(Food and Agriculture Organization of the United Nations,FAO)和国际原子能机构(International Atomic Energy Agency,IAEA)的突变品种数据库(Mutant Variety Database,MVD)统计,我国采用诱变手段已育成800 多个不同类型的植物新品种,约占世界各国育成品种总数的25%[13]。 近几十年来,随着我国航天科学技术的快速发展,特别是返回式科学实验卫星成功发射和回收,航天育种技术研究得到迅速发展,利用空间独有的特殊环境可以使小麦及其他植物基因发生变异,进而产生自然界稀有的或采用常规方法较难获得的新型突变体[14-16]。 目前,国内外对航天育种技术开展了广泛研究,本文主要阐述了航天育种的概念、发展历程以及所取得的成就,并重点介绍了近年来航天诱变对小麦的生物学效应研究,旨在为我国小麦航天育种研究提供一定的理论参考。
1 航天育种及其发展
1.1 航天育种
航天育种又称为空间诱变育种,是将农作物、经济作物、花卉等植物干种子或组织搭载返回式飞行器送到太空,利用太空中各种射线、近地强磁场、微重力等地面无法模拟的特殊空间条件综合作用打破基因连锁,促进优异基因重组,促使基因变异,进而使其性状发生改变,然后经过地面人工选育出植物新品种、优异育种材料以及特色基因资源材料的育种方法[17-18]。
1.2 航天育种的发展
航天育种研究依赖于航天飞行器技术的发展水平,目前世界上仅有美国、俄罗斯和中国独立掌握了该项技术,且我国是世界上唯一发射过航天育种专用卫星的国家[15,17]。 目前,已有多个国家和地区开展植物航天诱变育种研究,航天诱变育种技术已被FAO 和IAEA 等国际权威机构认定为是一种新型的诱变技术。
1.2.1 国外航天育种概况 由于航天育种对航天飞行器技术的要求较高,国际上早期开展航天育种研究工作主要集中于美国和俄罗斯。 如前苏联(俄罗斯)在19 世纪50年代就开始开展空间环境对植物生长发育影响的研究,主要观察植物在太空条件下的生长情况,同时对一些重要生理生化指标进行测定[19];20 世纪90年代在空间搭载种子在地面的萌发、生长发育等方面进行了研究[20]。 之后,Levinskikh 等[21-23]在空间站内模拟地面环境种植超矮秆小麦,并对其发芽情况、茎长、小穗数、穗粒数等农艺性状进行调查和分析,探讨了航天搭载对空间站超矮秆小麦生长发育的影响。美国科学家将大麦、小麦、大豆等植物在航天飞行器和模拟空间实验室内进行种植观察,发现这些植物在微重力、强辐射、超真空的条件下能正常生长,且有些植物的蛋白质含量得到显著增加[24]。 近年来,日本、印度等国家也相继开展了空间植物的研究,已培育了近千种太空植物,航天育种技术已成为很多国家培育优良植物新品种和育种材料、创制特色基因资源的有效途径之一[13]。
1.2.2 国内航天育种的发展 1987年,我国首次利用返回式卫星将第一批农作物种子带到太空,开启了航天诱变育种研究,经过几十年的发展,我国航天诱变技术在育种应用过程中取得了大量突破性的成果[15,25],其中“实践八号”育种专用卫星对推动我国航天育种研究发挥了重要作用[26]。 随着我国首颗微重力科学实验卫星“实践十号”成功发射,相关单位开展了搭载植物后续地面新品种选育、优异材料和特色基因资源的鉴定、筛选、利用等工作,保持了我国航天育种工作的持续性。 截至目前,我国共进行了20 多次航天育种搭载试验,搭载品种约1 200 种,先后选育出小麦、水稻、玉米、大豆、油菜、棉花、花生、芝麻等多个作物新品种,为我国农业发展做出了巨大的贡献[13]。
2 小麦航天诱变的生物学效应
大量研究表明,航天诱变能对小麦生物学效应产生重要影响。 如吕金印等[27]对不同小麦品种进行卫星搭载处理,研究SP2的主要农艺性状、光合速率、叶绿素含量的变化,田间观察发现12 份矮秆、高秆、早熟以及矮秆大穗的突变株系,表明卫星搭载能够引起较高的突变频率;王美芳等[28]研究发现小麦种子经航天诱变后能引起后代的广泛变异,SP3的株高、叶色、穗长、产量性状、品质性状的变异尤为突出,且这些突变性状在后代中会产生分离和遗传,多数分离类型通过后代选择可以使其趋于稳定。 成健[29]研究空间环境对冬小麦陕253 和西农1043 生长发育的影响及诱变效应,发现SP7突变株系的株高、穗长、单穗重、穗粒数、穗粒重、千粒重等农艺性状具有不同程度的变异,且均能够稳定遗传,其中陕253 和西农1043 的SP7少数突变株系在株高、千粒重、产量等性状与其对照的差异达到极显著水平。 但郭怡璠等[30]对春小麦品种龙麦26 和克丰10 号进行卫星搭载后,经地面种植后选育出变异品系,对其株高、穗长、小穗数、千粒重、容重以及产量等农艺性状进行调查后发现,变异品系较原品种在农艺性状上均有所变异,但差异均不显著。 综上,不同小麦品种对空间环境的敏感性存在差异,其诱变效果和性状变异的方向也存在着明显的差异。
3 小麦航天诱变的遗传机制
随着高通量测序、基因芯片、蛋白质组学以及分子标记技术的快速发展和应用,航天诱变研究已从表型鉴定、评价和利用,逐步发展到对诱变后代进行分子鉴定、变异特征和遗传规律分析、主效基因克隆和功能解析等诱变机理研究。 如Visscher 等[31]利用基因芯片技术对SP4小麦叶片与对照叶片的每个基因转录本丰度进行t 检验分析,发现二者小麦基因家族基因均未表现出显著性差异,表明经过太空搭载后小麦的基因表达模式不会出现显著的、可遗传的变化。 李鹏等[32]研究航天搭载对小麦醇溶蛋白和基因组DNA 变异的影响,发现航天搭载诱变对醇溶蛋白不同区域的敏感程度不同,以ε 区最为敏感,产生的变异最多,其次是β 区,而α 区和γ 区醇溶蛋白变异最小,航天诱变SP3材料经SRAP 标记分析后发现有多条多态性条带,且与对照间的遗传相似系数为0.821 ~0.937,说明航天搭载诱发了小麦基因组DNA 的变异。 郭怡璠等[30]对航天诱变材料基因组DNA 的多态性进行分析,发现B组染色体上的基因突变频率最高,A 组染色体的基因突变频率最低,D 组染色体上的基因突变频率介于二者之间。 而此前对“实践八号”卫星搭载获得的240个SP2突变体进行标记分析时发现,在A 组染色体上存在较高的基因突变频率[33],推测空间搭载导致小麦DNA 水平的变化是随机发生的,且对小麦基因组产生的诱变主要发生在重复序列区域,基因表达区域相对较少。 蒋云等[34]利用ISSR 技术研究空间环境诱变对小麦分子水平变异的影响,发现不同小麦基因型对空间环境诱变的敏感性存在明显的差异,与野生型相比,突变系在多个位点发生了DNA 水平上的改变,说明空间环境诱变引起了小麦分子水平的变异。
大量的研究表明,航天诱变不仅能够引发小麦在DNA 水平上发生变异(碱基替换、缺失、易位等),而且还能改变蛋白质表达。 如航天搭载小麦SP5突变株系的麦谷蛋白亚基变异类型较丰富,且变异频率较高,高分子麦谷蛋白亚基变异主要发生在GluB1 与GluD1 位点,同时发现所有SP5突变株系的总蛋白质含量均低于野生型,且在部分株系中发现与品质性状关系密切的谷蛋白大聚合体含量明显升高[29,35]。 此外,利用航天诱变技术获得丰富、优异的育种材料的同时,也获得了一批珍贵的基因资源。 如宋素洁等[36]通过航天诱变得到一个小麦叶绿素缺失的突变体Mt6172,并发现此突变体存在白化致死现象,推测可能是由于光合作用主要蛋白复合体缺失、叶绿体抗氧化能力下降或叶绿体RNA 转录后编辑途径受阻等原因造成的。 之后,Shi 等[37]采用转录组和叶绿体蛋白质组的方法对该突变体和野生型的差异表达基因和蛋白质进行鉴定分析,发现二者的大多数差异表达基因参与叶绿体发育和叶绿素的合成,进一步分析表明绝大多数转录蛋白质与光合作用和叶绿体合成相关,而光合蛋白编码基因的表达水平明显下降,这也为深入研究小麦叶绿体发育机制奠定了一定的基础。 Xiong 等[38]利用航天诱变技术并结合苗期耐盐性鉴定,筛选获得了1 个小麦耐盐突变体st1,发现该突变体在经过高盐处理后其发芽率与野生型相比显著提高,对野生型和SP5航天诱变突变体st1 进行转录组突变位点分析,发现航天诱变产生大量的单核苷酸多态性,而仅有少数的大片段缺失和插入。 进一步分析盐胁迫响应的野生型和st1转录组表达谱,发现在氧化还原过程中淀粉和蔗糖代谢、半乳糖代谢及亚油酸代谢调节在突变体st1 耐盐过程中发挥重要作用。 结合60Co-γ 射线、EMS 等其他诱变因素诱发的转录组序列变异情况,可揭示航天诱变突变体的转录组序列变异特征,为深入研究小麦航天诱变机理提供新思路和新方法。 利用航天诱变技术创制这些丰富、独具特色的基因资源材料也为进一步研究航天诱变机理和深入解析小麦重要经济性状形成的遗传机制奠定了基础。
4 我国小麦航天育种取得的成就
4.1 航天诱变小麦新品种的选育
航天育种技术与传统杂交育种相结合能快速有效地选育高产、优质、多抗等性状优良的小麦新品种。 河南省农业科学院小麦研究所是较早开展我国航天诱变育种工作的单位之一,在20 世纪90年代曾育成太空1 号小麦,该品系较原品种穗数和粒重均有提高,增产5%以上[39-40]。 之后,通过航天诱变技术选育了太空5号[39]、太空6 号[40]、郑麦3596[41]等小麦新品种。 其中,弱春性、优质、高产小麦新品种太空5 号是我国采用航天诱变技术选育的中早熟小麦品种,该品种达到国家优质弱筋小麦标准,适宜制作糕点、饼干等食品[39]。 太空6 号为半冬性、高产、分蘖力强、抗寒性好、品质较好的品种,其中面粉白度和淀粉糊化特性符合我国优质面条标准[40]。 针对优质强筋品种郑麦366 千粒重偏低、纹枯病抗性较差的问题,经过卫星搭载连续多年系谱法选择育成了优质、强筋、矮秆、抗倒小麦新品种郑麦3596,其综合品质指标达到了国家强筋小麦标准[41]。 河南省科学院同位素研究所在小麦航天与辐射育种方面也取得了丰硕的成果,其中以豫麦57 为材料,通过辐射诱变和太空搭载培育出高产、稳产、抗逆和广适性的小麦新品种富麦2008,该品种与原始亲本相比,黑胚率显著降低,综合抗性、落黄等性状得到明显改善,2005-2007年在黄淮南片累计示范种植8 万hm2,平均单产达到8 250.0 kg·hm-2,且出现多个9 750.0~10 500.0 kg·hm-2的高产典型[42]。 黑龙江省农业科学院作物育种所利用卫星搭载已先后培育出龙辐麦l5、龙辐麦18、龙辐麦19 等多个小麦新品种。 其中,龙辐麦l5 是我国第一个审定推广的航天诱变春小麦新品种,两年全省区域试验比对照品种新克旱9 号平均增产8.13%,达到极显著水平[43]。 龙辐麦18 由龙94-4083 纯系种子经卫星搭载后系统选育而成,该品种与原始亲本相比,生育期长2 d,株高高5 cm 左右,其产量三要素(成穗数、穗粒数和千粒重)也有所改善,较亲本增产5.3%以上,达到显著水平[44]。此外,山东、天津、甘肃、陕西等省份也都在早期广泛开展了小麦航天诱变育种研究,并取得丰硕的成果[45-46]。 通过航天诱变技术培育的部分小麦品种的信息详见表1。

表1 我国航天诱变技术培育的小麦品种(不完全统计)Table 1 Wheat varieties cultivated by space-flight mutagenesis techniques in China(incomplete statistics)
4.2 优异新材料的育种利用
目前我国现有小麦种质资源库遗传资源日益狭窄,严重阻碍了小麦遗传改良[47-48]。 利用航天诱变技术创制的优异的育种材料,能丰富我国小麦育种资源,满足小麦遗传育种的需求。 我国烟台市农业科学研究所20 世纪90年代初期将高产小麦品种鲁麦14 进行太空搭载诱变处理,创制出丰产、优质、抗病新种质烟航选2 号[49],之后,以烟航选2 号为亲本先后培育了烟农5160、烟农999 等优异小麦新品种。 山东省农业科学院原子能农业应用研究所从1994年开始,利用返回式卫星搭载创新小麦种质资源已选出一大批具有特异性状的突变体,其中早熟突变体SP121 和济南16 的诱变系940004 进行常规杂交后系统选育出高产、优质小麦新品种鲁原301[50];利用航天突变体9940168 和济麦19 进行常规杂交后系统选育出高产、优质小麦新品种鲁原502,并于2011年通过国家审定后,又陆续通过山东、安徽、江苏等省份审定或认定,在山东省累计推广面积达到167 万hm2,已成为山东省小麦种植第一大品种。 中国农业科学院作物科学研究所利用航天诱变早熟突变体SP121 与农大3338 杂交F1幼穗经γ 射线辐照后,连续多年种植从中选育出高产、抗旱、节水小麦新品种航麦901[51];而利用航天搭载后代材料选出的大穗早熟突变体G55 和轮选987 配制杂交组合,并结合基因诱变、花药培养等现代分子育种技术选育出高产、多抗小麦新品种航麦247,之后又利用航天搭载后代材料选出的优系SPLM2 和轮选987 配制杂交组合系统选育出高产、大穗小麦新品种航麦2566。 黑龙江省农业科学院作物育种研究所用九三3u90×九三少杂交后代纯系材料经航天诱变的SP4与优质亲本龙麦26 杂交,按系谱法选育而成高产、优质、抗病小麦新品种龙辐麦19,于2011年通过黑龙江省农作物品种审定委员会审定并推广[52]。 河北省农林科学院利用“实践八号”航天育种专用卫星搭载冬小麦轮选987,从中选育出矮秆、多穗、白粒突变系衡辐9103,在2012年国家预试中较对照良星99 增产3.17%[53]。 河南省科学院同位素研究所利用航天搭载诱变筛选出周麦18 的高产、矮秆突变系豫同194 和矮抗58 的矮秆、早熟、高产、配合力强的突变系豫同198。 其中豫同194 与豫麦34-6 配制杂交组合,系谱法选育高产、稳产小麦新品种郑品麦24,目前已通过第八届河南省主要农作物品种审定委员会审定,豫同198 与周麦18 配制杂交组合后系统选育出小麦新品种豫丰11,该品种已通过第四届国家品种审定委员审定。 同时,豫同198 与开麦18 配制杂交组合后系统选育郑品麦22,目前正参加2018年度黄淮南片生产试验。 以航天诱变技术筛选的优异育种材料为亲本杂交选育的部分小麦品种的信息详见表2。
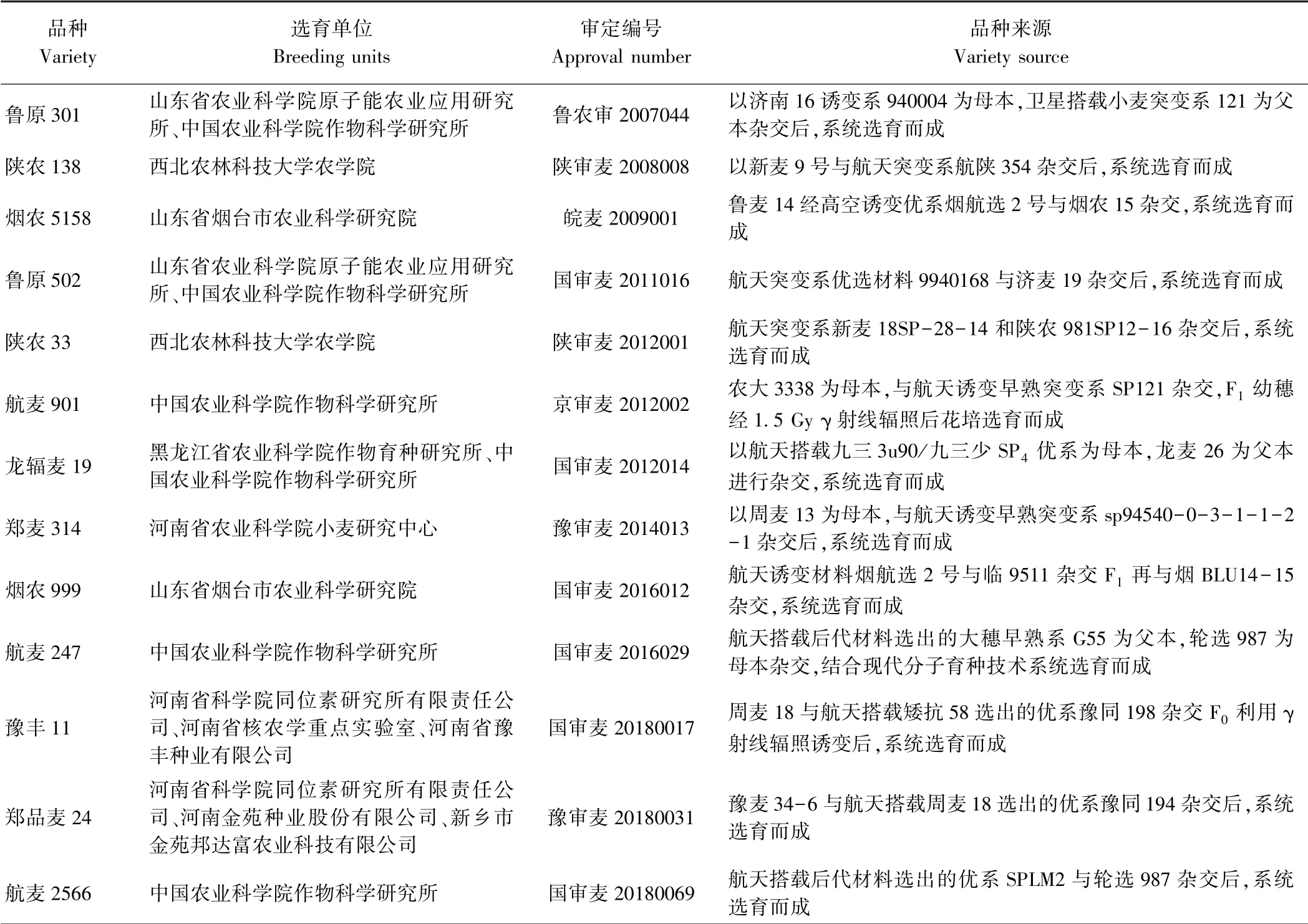
表2 我国利用航天诱变突变体杂交选育的小麦品种(不完全统计)Table 2 Hybrid breeding wheat varieties by space mutation mutant in China(incomplete statistics)
5 展望
应用航天诱变育种时应更加注重搭载材料的纯度,尽量杜绝剩余遗传变异带来的影响,同时地面种植时设置相应的对照。 对搭载材料干种子进行切分,只保留胚乳部分,既保证了种子的活力,又降低了搭载成本,从而极大地提高航天搭载的效率。 此外,航天搭载处理后切分后的小麦种子出苗势普遍较弱,因此需要相应的精细整地、足墒下种、适当浅播、加强苗期管理等配套栽培技术,以确保搭载材料的成活率,为遗传变异提供更可能多的选择群体。 近年来,关于我国小麦航天诱变育种技术基础研究和加强国际间诱变育种的合作与交流等方面取得了进展,如探讨了空间环境对小麦损伤效应和诱变效应[54]、太空环境与60Co-γ射线对小麦的不同诱变效应[55]以及利用转录组和蛋白质组的方法揭示航天诱变突变体的转录组和蛋白质组序列变异特征等[37-38],但研究的深度和广度与其他育种方法相比尚有较大差距。 随着小麦基因组研究计划的实施,小麦基因组、转录组和蛋白质组数据库的逐渐完善,利用高通量测序、转录组以及蛋白质组技术对航天诱变获得的变异材料开展系统性研究以阐明航天诱变的遗传和分子机理将是今后研究的重点。
综上所述,航天诱变育种除了具有育种周期短、有益突变频率高、目标性状稳定快、生理损伤轻、能快速打破基因连锁、促进优异基因聚合等优点外,还能与传统育种技术、细胞工程技术、分子标记育种技术相结合,多种技术及学科的协同互作,将促进小麦航天诱变育种研究得到进一步发展和提升,在今后小麦产量的提高、品质的提升、综合抗性增强等方面发挥重要的作用。 但影响航天诱变效果的因素极为复杂,既有来自空间宇宙射线、微重力、高真空、交变磁场等外在因素,也有异源多倍体、基因组庞大等自身因素,因此小麦航天诱变机理的研究难度极大。 航天诱变技术在我国小麦育种方面虽已取得了巨大的成就,但有关小麦航天诱变育种的理论、方法和高效育种技术体系还有待进一步完善。 此外,运载火箭和航天飞行器的研制、发射以及回收等对技术、资金的要求极高,因此航天诱变方法相对其他理化诱变方法而言,其成本较高,此外,航天诱变获得的性状变异范围较窄,给后期的田间选择和室内鉴定带来一定的难度。 目前,我国小麦航天诱变育种的应用研究已处于世界领先水平,但航天诱变育种机理、提高诱变效率等基础理论研究仍十分薄弱,从细胞学、生理生化和分子生物学等方面对空间诱变生物学机理的研究相对较少。 因此,解析空间诱变的遗传规律,加强对空间环境的地面模拟研究,对提高我国小麦航天诱变育种水平至关重要。
猜你喜欢
上海农业科技(2022年3期)2022-06-11
四川蚕业(2021年2期)2021-03-09
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
河北果树(2020年2期)2020-05-25
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
天津医科大学学报(2015年2期)2015-12-22
百科知识(2015年18期)2015-09-10
山东医药(2015年40期)2015-02-28
