
日本血吸虫虫卵分泌物小RNA的高通量测序
2019-02-12 03:30徐桂娜何雪梅周晓蓉曾凡胜秦志强
热带病与寄生虫学 2019年4期
徐桂娜 何雪梅 周晓蓉 曾凡胜 秦志强
血吸虫病是严重危害群众身体健康,阻碍经济社会发展的寄生虫病[1]。随着我国血吸虫病防治工作的不断深入,当前我国血吸虫病已呈低度流行态势[2]。但目前经典沿用的诊断方法都存在不同程度的缺陷,越来越难以满足当前防治的需求[3]。因此,现阶段迫切需要建立与发展精准的检测方法用于我国血吸虫病防治[4]。
微小RNA(microRNA, miRNA)是一类长18~30 nt高度保守的内源性单链非编码小RNA(small RNA,smRNA)分子[5],通过结合互补mRNA调节基因表达,参与细胞生长、分化、增殖与凋亡、新陈代谢等多种生命活动[6]。miRNA在血吸虫生长发育、寄生及致病机制中可能具有重要作用[7]。因此,对miRNA在日本血吸虫病中的深入研究可为药物靶点预测、疫苗研制及诊断开辟新的思路。近年来,围绕血吸虫miRNA的鉴定、靶基因识别及诊断展开了较多研究。目前研究miRNA表达的主要方法有Real-Time PCR、miRNA芯片以及第二代测序[8]。本文通过构建日本血吸虫虫卵分泌物(egg secreted product, ESP)的小RNA文库,应用Illumina NextSeq 500第二代高通量测序技术对小RNA文库进行测序,再结合生物信息学分析方法,快速鉴定保守的miRNA,筛选出差异表达的miRNA,为下一步验证血吸虫感染患者血液中的miRNA靶标并为发展基于miRNA的分子诊断方法奠定基础。
材料与方法
一、日本血吸虫ESP制备
参照朱蓉[9]等报道的方法制备日本血吸虫虫卵。新鲜的日本血吸虫虫卵用0.9%生理盐水和RPMI1640培养基分别漂洗3次,用RPMI1640培养基于37℃、5%CO2培养12 h后,吸取培养上清,1200×g、4℃离心10 min,收集上清,即为虫卵分泌物(ESP),虫卵ESP用0.45 μm的无菌滤器过滤后置-80℃保存备用。
二、ESP总RNA制备
使用Trizol试剂(Invitrogen,美国)提取日本血吸虫虫卵ESP总RNA,利用日本Takara提供的专用试剂37℃孵育30 min以清除残留的基因组DNA,使Oligotex mRNA MidiKit(Qiagen,德国)分离纯化样品mRNA。RNA的质量和含量通过使用紫外可见分光光度计(Nanodrop ND-1000,LabTech,美国)检测吸光度260 nm/280 nm(A260/A280)进行测定,完整性通过1.5%(w/v)凝胶电泳进行检测。
三、测序文库构建和RNA高通量测序
按照小RNA测序文库构建流程对纯化后的日本血吸虫ESP总RNA进行3′端接头连接,5′端接头连接、反转录、扩增,cDNA文库大小选择,纯化等步骤,完成测序样本文库构建。构建好的文库利用荧光染料法(Picogreen)和荧光光度计方法定量文库,并用Agilent 2100 Bioanalyzer(美国)和ABI StepOnePlus Real Time PCR System(美国)进行质检,将每个质检检测合格的样品稀释至终浓度10 nmol/L后,交由上海派森诺生物技术有限公司,使Illumina NextSeq 500测序仪(美国)进行测序。
四、原始数据预处理、数据过滤及长度分布统计
应用质控工具fastx(fastx_toolkit-0.0.13.2)对测序原始数据进行预处理,去除接头序列以及低质量序列(包括模糊碱基N,碱基质量小于20以及长度小于18 nt的序列),得到可供后续分析的序列,并进行处理结果统计及长度分布统计。
五、比对注释,鉴定已知miRNA
应用CLC genomics_workbench 5.5商业软件将预处理后的序列与最新Sanger miRBase数据库比对,比对过程不允许有碱基错配。另与其他非编码数据库ncRNA、piRNA和Rfam数据库比对,允许与目标序列有2个碱基错配、允许比目标序列两端各缩短或延长2个碱基。
六、miRNA表达水平
采用DEGseqR语言包结合perl脚本将ESP按照分组情况进行miRNA表达量的比较分析。使用标准化数值(Transcripts per million,TPM)进行差异分析。
结 果
一、总RNA提取质量评估
RNA的质量和含量通过使用Nanodrop ND1000(LabTech,美国)检测,吸光度260 nm/280 nm(A260/A280)的值为2.33,完整性通过1.5%(w/v)凝胶电泳进行检测,RNA完整性计数RIN(RNA Integrity Number)为2.4(见图1)。
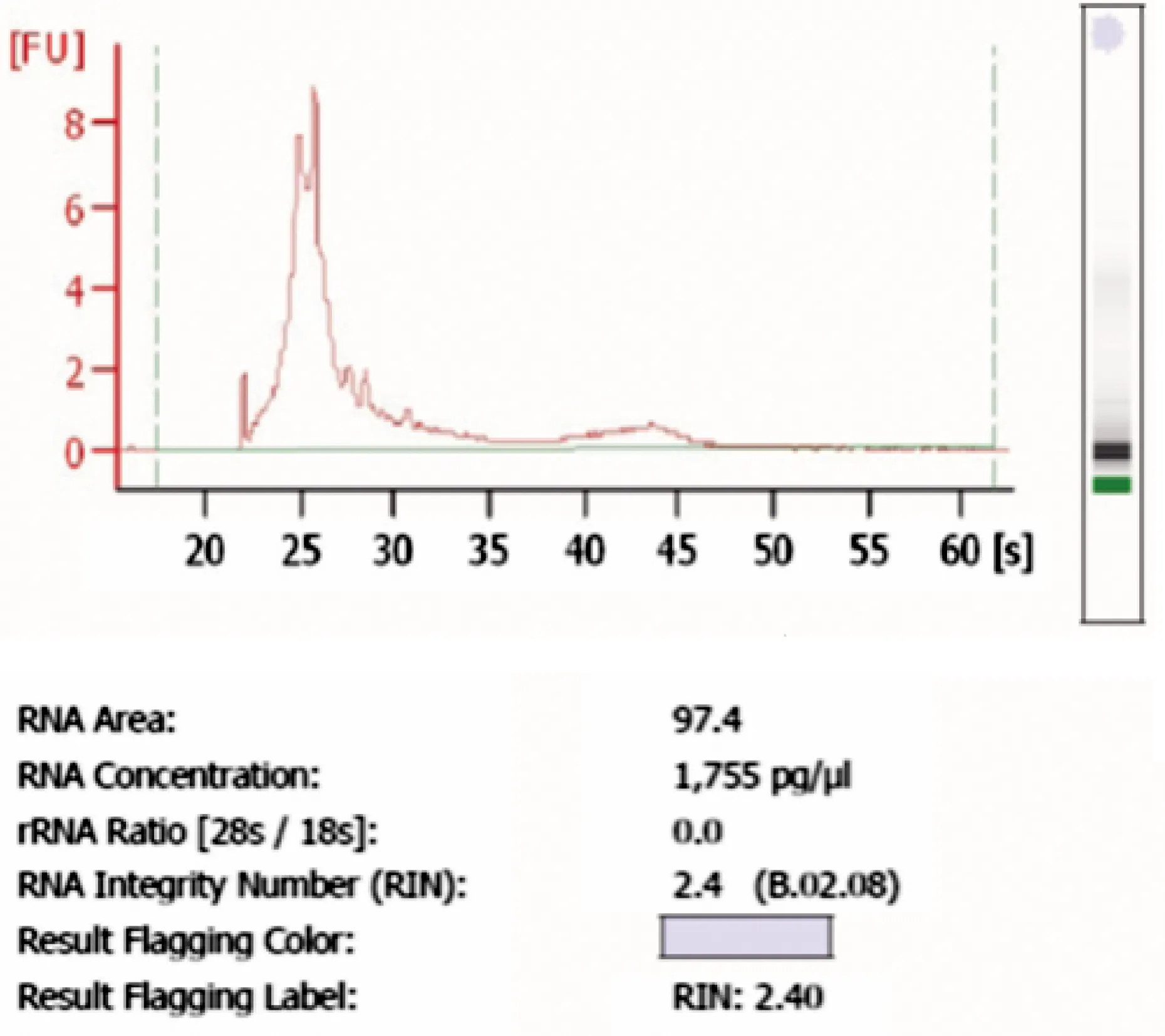
图1 总RNA提取质量分析
二、原始数据预处理、数据过滤
本研究利用IlluminaNextSeq 500测序平台对血吸虫ESP进行测序分析获得原始阅读序列读数(Raw Reads)为23 348 390条。应用fastx(fastx_toolkit-0.0.13.2)对日本血吸虫虫卵ESP测序,原始序列数(reads)进行预处理,去除接头序列以及低质量序列(包括模糊碱基N,碱基质量小于20以及长度小于18 nt的序列),最终获得ESP过滤后的序列读数为13 190 761条。去掉重复序列前ESP序列长度分布统计在30 nt处出现最高峰;去重后ESP序列长度分布统计主要分布在26~30 nt处(见图2)。

横轴为不同的序列长度,纵轴为对应长度的序列丰度(*10000)
三、比对注释及miRNA表达
将测序所得小 RNAs序列进行比对注释,其中有注释的小 RNA为84 735条,约占12.6%。将构建文库中样品表达的miRNA与最新Sanger miRBase数据库中的miRNA比对,ESP文库中有270条序列可与日本血吸虫的miRBase数据库比对上,已知的成熟miRNA有12个,未发现有新miRNA。miRNA表达量分析显示sja-miR-3488为表达量最高的miRNA,sja-miR-36-3p和sja-miR-71a表达量也较高,3种miRNA表达量占91.8%(见表1)。

表1 miRNA表达定量分析
讨 论
miRNA在哺乳动物血浆、血清、尿液等体液中稳定存在[10,11],近年来已相继发现循环miRNA可作癌症的生物标志物,是研究者高度关注的新诊断靶点[12~14]。miRNA亦用于诊断多种其他感染性疾病的生物标志物,比如肺结核[15,16]、脓毒病[17,18]、病毒性肝炎[19,20]等。近年来,人们在血吸虫感染宿主的血液[21,22]、粪便[23,24]中检测出了血吸虫虫体、虫卵来源的循环DNA,为血吸虫病的核酸诊断研究开辟了新的途径。miRNA在日本血吸虫虫卵和虫体有特异性表达[25],有可能成为特定生物标志物。日本血吸虫虫卵sja-miR-71b-5p、sja-miR-71、sja-miR-36-3p高表达[25],sja-miR-125b在成虫呈显著表达[26],提示上述miRNAs分别具有作为虫卵和成虫阶段特异标志物的潜力。Hoy等[27]在感染曼氏血吸虫小鼠血清中鉴别出宿主源性和寄生虫源性的miRNA,并在感染患者血清中检测出的三种寄生虫源性miRNAs,即miR-277、miR-3479-3p和miR-bantam,上述三种miRNAs可作为血吸虫感染的新标志物。Cai等[28]发现寄生虫源性的特异循环miRNAs sja-miR-277和sja-miR-3479-3p是诊断日本血吸虫病的潜力靶标。以上研究显示血吸虫病患者血清或血浆中出现特异的miRNA,且具有稳定性高、结合灵敏度高和检测特异的特点。虫卵作为主要致病因素可促使血吸虫感染的发生,并且逃避或操纵宿主免疫反应,因此对血吸虫虫卵ESP miRNA的研究有助于发现新的干预目标[25,29]。为鉴定日本血吸虫ESP miRNAs,本研究基于Illumina高通量测序技术对日本血吸虫ESP miRNA进行了测定,获得了13 190 761个可用的过滤后的序列数,经过对过滤后的数据进行比对注释,获得12个已知miRNAs,未发现有新miRNA,其中sja-miR-3488表达量最高,sja-miR-36-3p和sja-miR-71a表达量也较高,这三种miRNAs表达量占91.8%。而血吸虫虫卵中sja-miR-71b-65p、sja-miR-71、sja-miR-1、sja-miR-36-3P、sja-miR-124-3p表达最高,占86%,说明特定miRNA家族在寄生虫的不同发育阶段都有特征性的偏倚表达[25]。sja-miR-36-3p在血吸虫胚胎发育中起重要作用,同时也可调节血吸虫尾蚴和肺期童虫发育;sja-miR-71家族成员在虫卵阶段发挥重要的调节作用[25]。
综上,通过对血吸虫ESP中miRNAs的鉴定,尤其是sja-miR-3488、sja-miR-36-3p和sja- miR-71a可能在血吸虫病虫卵致病过程中具有重要作用,它们是否存在于血吸虫感染宿主血清中,值得进一步验证和阐明其功能以寻找新的血吸虫病诊断标志物。
猜你喜欢
今日健康(2022年3期)2022-11-21
电影故事(2021年1期)2021-08-16
科学之谜(2021年2期)2021-04-25
——以江苏省为例
云南档案(2020年7期)2020-12-06
科学导报(2020年54期)2020-09-09
学习与科普(2019年13期)2019-09-10
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
特别健康·下半月(2018年3期)2018-04-23
家庭用药(2018年3期)2018-03-23
