
基于RNA-seq分析黄曲霉毒素B1诱导大鼠肝癌的可变剪接事件
2018-12-04 05:16谭妮曹骥杨春郑海平欧超
中国癌症防治杂志 2018年5期
谭妮曹骥杨春郑海平欧超
肝癌是常见的恶性肿瘤,全球5%~28%的肝细胞癌发病可归因于黄曲霉毒素暴露[1-2]。黄曲霉毒素暴露可增加慢性乙型肝炎病毒(HBV)感染患者罹患肝癌的风险[3-5]。黄曲霉毒素 B1(Aflatoxin B1,AFB1)和乙型肝炎病毒感染对肝癌发生发展有相互协作作用,但AFB1诱发肝癌的具体机制尚未清楚。可变剪接的总体功能是增加mRNA的多样性,可通过调节信号通路中的一些关键基因表达,在动物生长、发育、生理代谢过程中发挥重要调控作用。可变剪接的改变大多由点突变引起[6],可产生多种蛋白质同种型,对恶性肿瘤发展至关重要[7-9],其与恶性肿瘤的关系亦成为研究热点之一。本研究通过AFB1诱发大鼠肝癌模型,采用RNA-seq高通量测序探讨大鼠肝脏组织的可变剪接事件,以期阐明AFB1诱发大鼠致肝癌的可能机制。
1 材料与方法
1.1 动物分组
90只Wistar雄性大鼠由广西医科大学动物实验中心提供,4周龄,体重40~60 g。单独饲养于不锈钢笼,室温(23±1)℃,相对湿度(70±10)%。AFB1购自Sigma公司。经4周适应期后,将大鼠随机分为AFB1组60只和对照组30只。AFB1组大鼠腹膜内注射AFB1,剂量:4~7 周、9~12 周,3 次/周,每次 200 μg/kg;14~17 周、19~22 周、24~27 周、26~32 周,2 次/周,每次 100 μg/kg;34~37 周、39~42 周、44~47 周、49~52周,1次/周,每次100 μg/kg。对照组以正常饲料喂养。在第70周脱颈处死大鼠,取肝组织并置于液氮冷冻,用于提取肝组织RNA。本实验经实验动物伦理委员会审核。
1.2 RNA提取
采用传统TRIzol(购自美国Invitrogen公司)提取RNA,130~150 mg肝组织用液氮研磨,在TRIzol裂解5 min,加氯仿静置 2 min,12 000 r/min 4°离心 15 min,取上层液体,加入等量异丙醇静置10 min,12 000 r/min 4°离心 15 min,75%无水乙醇清洗沉淀物 2次,12000r/min 4°离心15 min,取适量DEPC水溶解沉淀物。琼脂糖凝胶电泳检测RNA完整性,凝胶成像系统观察总RNA的5s rRNA、18s rRNA和28s rRNA条带,3条条带完整即证明总RNA提取较完整。BioPhotometer plus艾本德核酸蛋白测定仪检测OD值,OD260/OD280比值大于1.8,则说明制备的RNA较纯,无蛋白质污染。
1.3 RNA-seq测序
RNA-seq测序由华大基因科技有限公司完成,采用Illumina HiSeq 2000进行RNA测序,读取长度为90 pb。采用 FastQC工具(http://www.bioinformatics.babraham.ac.uk/projects/fastqc/)检查 RNA-seq测序数据的质量,Tophat2(v2.0.9)将序列比对至大鼠基因组(Rnor_5.0.75)进行序列比对,将读段映射到参考基因组进行剪接位点预测。
1.4 可变剪接事件鉴定
选择目前已知的具有多种可变剪接存在的332个参考基因。采用Cufflinks(v1.3.0)软件计算差异表达基因同种型水平的FPKM值,找出在不同大鼠肝组织中存在差异表达基因的同种型,并将其比对到DAVID数据库,获得GO分析生物过程(biological process,BP)功能富集。
2 结果
2.1 大鼠一般情况
至第70周,AFB1组大鼠存活42只,其中发生肝癌 31只(成癌组),未发生肝癌 11只(未成癌组),对照组30只大鼠均存活。
2.2 RNA-seq测序数据的质量
对照组、成癌组、未成癌组序列的5′端前12个位置都有碱基(AGCT)比例不均匀的情况,故将前12个碱基切除,将剩余的78 bp用于比对分析(图1)。3组大鼠组织中比对上的基因组序列占原始序列的比例均在85%以上,说明3组大鼠肝组织的测序质量较高,可用于后续分析。
图1 双端测序左侧和右侧序列碱基质量得分在不同位置上的分布
2.3 基因可变剪接差异分析结果
RNA-seq测序结果显示,成癌组与对照组共有37个基因的可变剪接模式存在差异,最显著的前15类GO分析生物过程显示,这些差异基因主要富集在刺激反应功能和细胞增殖调节功能上(图2A);未成癌组与对照组共有18个基因的可变剪接模式存在较大差异,差异基因主要富集在刺激反应功能上(图2B);成癌组与未成癌组有32个基因的可变剪接模式存在明显差异,差异基因主要富集在细胞增殖调节功能上(图2C)。
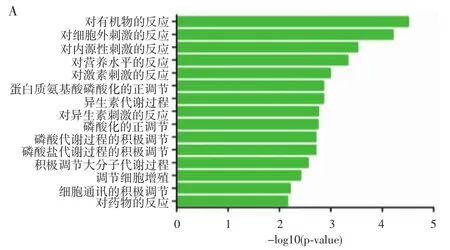

图2 各组差异表达基因最显著的前15类GO分析生物过程
2.4 成癌组中与细胞增殖调节有关的基因
分析成癌组细胞增殖调节相关基因的异常可变剪接模式,发现7个与细胞增殖调节相关的基因的可变剪接模式发生异常,分别是KIT配体(Kitlg)、自噬关键基因(Becn1)、小窝蛋白-1(Cav1)、组蛋白精氨酸甲基转移酶 1(Carm1)、胰岛素样生长因子 1(IgfI)、视黄酸受体 γ(Rarg)、转录因子 E2a(Tcfe2a),其中成癌组、未成癌组和对照组IgfI、Carm1、Tcfe2a基因可变剪接模式存在显著差异,见图3。

图3 各组大鼠肝组织中差异基因的可变剪接模式
3 讨论
RNA-seq可以准确检测基因的表达量,精确地识别基因序列变化,被广泛应用于检测动植物基因组中发生的可变剪接事件[10-13]。为了研究大鼠在黄曲霉毒素作用下发生的可变剪接差异,本研究通过RNA-seq对成癌组、未成癌组及正常对照组大鼠肝组织的基因表达进行检测并分析其可变剪接事件。可变剪接是一个过程,即由主要基因或者mRNA前体转录所产生的RNA的外显子以多种方式通过RNA剪切进行重连,由此产生不同的mRNA可能被翻译成不同的蛋白质异构体,因此一个基因可能编码多种蛋白质,可变剪接在调节基因表达和转录组多样性、蛋白质组多样性中起关键作用。本研究成癌组与对照组经比较发现可变剪接模式存在显著差异的基因主要富集在细胞增殖调节功能以及刺激反应功能中,而未成癌组与对照组差异表达基因只富集于刺激反应功能上,说明无论大鼠肝脏组织是否发生癌变,AFB1的加入均会导致大鼠肝脏细胞对AFB1这一外源物质产生应答反应,而且这一过程涉及部分基因的可变剪接体选择。值得注意的是,成癌组与未成癌组的差异表达基因富集于细胞增殖调节功能上,说明在AFB1作用下,大鼠是否发生肝癌病变可能主要与细胞增殖调节有关。
进一步研究参与AFB1诱发大鼠肝癌过程中细胞增殖调节的差异基因,发现共有7个差异基因参与了成癌组的细胞增殖调节过程,其中成癌组和未成癌组中IgfI基因、Carm1基因、Tcfe2a基因的可变剪接模式有明显差异。成癌组中Tcfe2a基因的两个同种型表达均高于对照组和未成癌组,说明大鼠在受到外源性AFB1刺激后,Tcfe2a基因的可变剪接模式改变可能与其致大鼠肝癌有关。Beck等[14]研究也表明Tcfe2a编码的两个可变剪接体E47和E12中,E47主要对细胞发育、增殖起作用。Matsuda等[15]研究发现 Carm1在细胞内的活跃程度较高,选择性剪接对转录调节有重要作用。Wang等[16]研究亦报道Carm1的可变剪接调节与甲基化有关。本研究中,Carm1基因的两个同种型在成癌组中的表达高于对照组和未成癌组,提示Carm1基因的可变剪接模式可能对AFB1诱发大鼠肝癌有重要作用。Mourmouras等[17]研究发现Igf1不同转录本的表达对膀胱癌的发生有重要作用。本研究发现Igf1基因同种型的表达在成癌组、未成癌组和对照组中亦存在差异,提示Igf1的可变剪接模式在AFB1诱导大鼠肝癌形成的过程发生改变,可能参与大鼠肝癌的发生。
本研究结果显示,IgfI、Carm1、Tcfe2a基因的可变剪接模式在大鼠不同肝组织中存在显著差异,其可变剪接改变在AFB1暴露后的肝癌形成过程中可能发挥重要作用,这一研究结果为AFB1致肝癌的机制研究提供了新思路。
猜你喜欢
保健医苑(2022年4期)2022-05-05
昆明医科大学学报(2021年4期)2021-07-23
世界科学技术-中医药现代化(2021年10期)2021-03-02
汽车维修与保养(2021年8期)2021-02-16
中国生殖健康(2020年4期)2021-01-18
中西医结合肝病杂志(2020年2期)2020-10-27
中国现代中药(2019年5期)2019-07-03
科海故事博览·下旬刊(2019年6期)2019-04-16
中国生殖健康(2018年4期)2018-11-06
癌变·畸变·突变(2015年4期)2015-02-27
