
延安市人工刺槐林光合生理生态特性与生态效应研究
2018-10-30 10:35:06冯晓东
上海农业学报 2018年5期
李 萍,邓 凯,冯晓东
(延安大学生命科学学院,延安716000)
植物的光合作用不仅受内部各种生理因子的影响,还与其生境密切相关,研究植物光合生理生态特性可以揭示其生态适应机制。刺槐(Robiniapseudoacacia)因其根系比较发达、生长迅速、耐干旱瘠薄,而被选为黄土丘陵区的主要造林树种[1],一般应用于沟壑治理、矿区复垦、农田防护以及水利建设护坝工程中,对改善生态环境,防止水土流失,吸纳大气中的粉尘、净化空气,乃至在调节黄河水文状况中发挥着重要的作用[2-3]。近几年对刺槐的研究主要侧重在人工林碳、氮储量和刺槐林土壤生态化学等[4-13]方面,而对刺槐的光合生理方面还需进一步研究。本研究以延安市宝塔山上的刺槐为试验材料,通过植物光合作用的光响应曲线得到生理参数,摸清刺槐的光合特性,并在测定其夏季光合速率、蒸腾速率的基础上,进行了关于刺槐的固碳释氧、降温增湿的生态效应的量化研究。本研究主要目的在于为延安市刺槐的生理生态学研究和人工林主要树种生态效益、适应性评价提供理论基础和数据支持,也为指导陕北地区刺槐人工林建设提供依据。
1 材料与方法
1.1 研究区概况
宝塔山刺槐林,树龄19年左右,由全国各个工会义务种植。宝塔山坐落于延安城东南方,海拔1 135.5 m,属于暖温带半湿润气候区,全年气候干旱且受季风环流影响。降水比较集中,夏季多冰雹、暴雨。冬、春易旱,且有风沙或寒潮的侵袭,气象灾害较频繁。本研究选取延安市地区具有代表性的植物刺槐为试材,随机选取20 m×20 m样地,在样地任意选取长势一致的5株,并做标记进行长期监测。
1.2 方法
1.2.1 光合速率、蒸腾速率的测定
2017年8月份选择晴朗、无风的天气,利用GFS-3000便携式光合仪(德国)开路系统(空气流速为750 μmol·s-1)测定刺槐的净光合速率(A,μmol·m-2·s-1)和蒸腾速率(E,mmol·m-2·s-1),并同步测定光合有效辐射(PAR,μmol·m-2·s-1)、气温(T,℃)、叶温(T1,℃)、空气相对湿度(RH,%)、空气CO2浓度(Ca,μmol·mol-1)、胞间CO2浓度(Ci,μmol·mol-1)和气孔导度(Gs,mol·m-2·s-1)等相关参数值。选择刺槐的冠层阳面、半阴面和阴面中部枝条前端的生长状况相对一致的第3—5片叶片,做好标记。每组测定3株,重复3次,取平均值。9:00—19:00每隔1 h测定1次光合参数值。
根据测得的相关数据得出叶片气孔限制值(Ls)、水分利用率(WUE,μmol·mmol-1),计算公式如下:
Ls=1-CiCa
(1)
WUE=AE
(2)
1.2.2 光响应曲线测定
于9:30—11:30叶片经充分光诱导后,采用GFS-3000附带的LED-Panel红蓝光源控制叶室内的光强,光合有效辐射(PAR)设置为梯度式增长,分别为0 μmol·m-2·s-1、20 μmol·m-2·s-1、50 μmol·m-2·s-1、80 μmol·m-2·s-1、100 μmol·m-2·s-1、200 μmol·m-2·s-1、500 μmol·m-2·s-1、800 μmol·m-2·s-1、1 000 μmol·m-2·s-1、1 200 μmol·m-2·s-1、1 500 μmol·m-2·s-1、1 800 μmol·m-2·s-1、2 000 μmol·m-2·s-1,其他因子为自然状态下测定。根据公式拟合A-PAR曲线:
A=Amax(1-C0exp(-ΦIAmax))
(3)
式中:Amax表示最大光合速率,Φ是由光反应曲线的初始斜率得出的表观量子效率(AQY),I为光合有效辐射(PAR)。
1.2.3 固碳释氧量和释水吸热量的计算
设定一天的光合时间为12 h,估算单位面积叶片每天所吸收的 CO2量、释放的 O2量和蒸腾总量。
计算公式:A=日平均净光合速率(μmol·m-2·s-1)×光合时间(12 h)×3 600(s)
(4)
E=日平均蒸腾速率(mmol·m-2·s-1)×蒸腾时间(12 h)×3 600(s)
(5)
W(CO2)=A×44×10-6
(6)
W(O2)=A×32×10-6
(7)
W(H2O)=E×18×10-3
(8)
Q=W(H2O)×L×4.1868(1 cal=4.1868 J)
(9)
其中A(μmol·m-2·s-1)和E(mmol·m-2·s-1)分别表示测定日的同化总量和蒸腾总量,Q(J·m-2·d-1)表征单位面积叶片在测定日因释放水分而吸收的热量;W(CO2)(g·m-2·d-1)和W(O2)(g·m-2·d-1)表示单位面积叶片测定日固定 CO2的质量、释放 O2的质量;W(H2O) (g·m-2·d-1)表示单位面积叶片测定日释放水的质量,CO2、O2、H2O 的摩尔质量为分别为44 g·mol-1、32 g·mol-1、18 g·mol-1。
L代表蒸发潜热系数,表达式为:L=597-59T1[14-15],其中T1为蒸发面的温度(℃),0℃时的蒸发潜热为597 J。
1.3 数据分析
利用 Excel 2010进行绘图,SPSS 20.0进行统计分析,Pearson 方法进行相关分析,LSD 进行方差分析,并对外部不同环境生态因子和内部各种生理因子进行主成分分析和通径分析。
2 结果与分析
2.1 环境因子日变化
由图1可知,PAR日变化为“单峰”曲线,PAR和T在12:00达到峰值,分别为1 452.63 μmol·m-2·s-1、31.81 ℃,而RH和Ca反而最小,即44.71%和23.03 μmol·mol-1。这可能是因为随着PAR的加强,光合作用渐渐加强,固定的CO2逐渐增多,环境中的CO2降低下降;但随着温度升高,环境中的湿度便会逐渐下降。水气压饱和亏(VPD)出现两个峰值,分别在11:00(50.34 Pa·kPa-1)和15:00(39.83 Pa·kPa-1)。因此,各生态因子的变化可进一步表明试验当天为典型的晴朗天气,而且刺槐外部不同环境生态因子之间相互影响。
2.2 T1、E、Gs、Ci的日变化
刺槐的T1的日变化呈“单峰”曲线,而且日变化规律与T的日变化基本一致,日平均温度为33.21℃(图1c)。
刺槐的蒸腾速率(E)的日变化呈“单峰”型,15:00达到最大值3.8 mmol·m-2·s-1,随后随着PAR的下降,T下降,RH升高,最终E逐渐下降,到19:00下降到一天中的最低值,为0.21 mmol·m-2·s-1(图1e)。
刺槐的气孔导度(Gs)受环境影响较大,随PAR的增强,T逐渐升高,E随之逐渐增大,植物为了更好地储存水分避免热害只能渐渐关闭气孔从而保护自身,因此从09:00开始Gs一直在降低,在18:00达到最低值7.33 mmol·m-2·s-1。Gs变化趋势较大,由11:00的177.2 mmol·m-2·s-1大幅度降低到12:00的39.4 mmol·m-2·s-1,12:00—17:00T逐渐下降,Gs变化趋势较平稳(图1a)。
刺槐的胞间CO2浓度变化曲线近似呈“W”,在09:00处于较高水平,为 400.67 μmol·mol-1,随后逐渐下降,10:00到达第一个低谷,为 225.33 μmol·mol-1,随后缓慢上升,11:00到达一个拐点,之后大幅度下降,12:00到达第二个低谷,也为一天中的最小值,随后逐渐上升,18:00达到最大值,为 429.13 μmol·mol-1(图1d)。
2.3 叶片净光合速率日变化
在8月份刺槐的净光合速率(A)日变化为典型的“双峰”曲线,光合“午休”现象比较明显(图1e)。从早上开始随着光强的增加,A逐渐上升,刺槐的A最大峰值出现在11:00,值为5.03 μmol·m-2·s-1。中午12:00左右,随着PAR达到最大值,温度升高,蒸腾速率加快,刺槐的A达到第一个低谷,为0.58 μmol·m-2·s-1。之后又随着PAR的降低,刺槐的A逐渐升高,在15:00出现第二个峰值,为2.22 μmol·m-2·s-1。接着又逐渐下降,在18:00达到一天的最低值,为0.02 μmol·m-2·s-1。
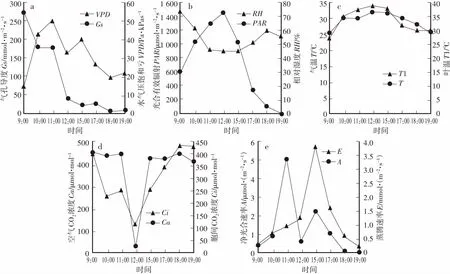
图1 主要生态因子日变化Fig.1 Diurnal changes of main ecological factors
2.4 净光合速率与相关生理生态因子的关系
2.4.1 相关分析
将刺槐的A分别与外部不同环境生态因子和内部各种生理因子进行 Pearson 系数双边检验,结果见表1。与A呈极显著正相关的是RH、Gs、Ci、Ca,与T1、T则显示为极显著负相关。刺槐的PAR与A不同,其与VPD、Ca、T1为极显著正相关,反而与RH呈极显著负相关。与RH呈显著正相关的有Gs、Ci、T,与VPD、Ca、T1呈负相关。刺槐的E只与Ci呈显著负相关。刺槐的VPD与T1、T呈极显著正相关,与Gs、Ci呈极显著负相关。对于刺槐的Gs而言,它与Ci呈极显著正相关,与T1、T呈极显著负相关。刺槐的Ci与Ca呈现出极显著正相关,与T1、T为极显著负相关。而刺槐的Ca和T1都与T呈极显著正相关,Ca与T1呈极显著负相关。

表1 刺槐净光合速率(A)与光合生理生态因子的相关系数矩阵
*、**分别为显著相关、极显著相关(P<0.05、P<0.01)
2.4.2 逐步回归分析
采用逐步多元回归分析法能有效地从众多外部环境生态因子和内部生理因子中挑出对Y(A)贡献最大的因子,并建立Y(A)与这些因子的最佳回归方程。用X1、X2、X3、X4、X5、X6、X7、X8分别代表刺槐的生理生态因子PAR、RH、E、VPD、Gs、Ci、Ca、Tl、T,对刺槐Y(A)以及各影响因子(X1—X8)分别进行逐步多元回归分析,得到回归方程为:Y=-8.484+0.021X5+0.282X8+0.002X7(R2=0.860),表明X5(Gs)、X7(Ca)、X8(T)是影响刺槐Y(A)的主要因子。
2.4.3 通径分析
为了更深入的明确各生理生态因子对A的直接和间接影响,对测定的结果进行通径分析,将回归方程Y的Gs、Ca、T对A的影响进行分解。如表2所示:关键因子对A的直接影响表现为:Gs(0.899)>T(-0.531)>Ca(0.228),Ca对A的表现为间接效应,且影响最小。通过分析比较各个间接通径系数发现,虽然Gs通过Ci对刺槐的光合速率Y产生一定负向的间接作用,但由于Gs与光合速率的相关系数和Gs通过T对刺槐光合速率Y的间接通径系数值较大,故Gs对光合作用Y的影响较大。T对刺槐光合速率Y的相关系数为-0.531,故T对光合速率Y的影响也较大。

表2 主要因子通径分析
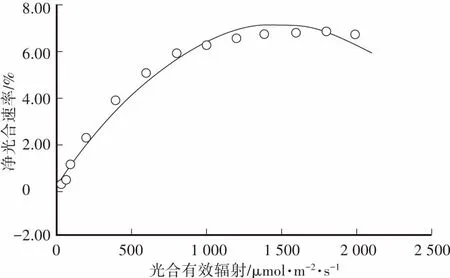
图2 刺槐的光响应曲线Fig.2 Light response curve

图3 刺槐的光-水分利用效率反应曲线Fig.3 Light-water use efficiency response curve
2.5 光强响应
刺槐的光响应曲线利用光响应实际观测数据和A-PAR公式拟合得到(图2)。刺槐的光响应曲线的拟合相关系数R2为 0.987,说明饱和光强和最大净光合速率(Amax)的拟合值与实际观测值都很接近。随着光合有效辐射的提高,刚开始刺槐的净光合速率上升较快,随后逐渐变缓,当到达一定限度时,即使再增加光照强度,净光合速率也不再增加,最终达到光饱和点(LSP),继续增加光照强度,多余的光能造成光合速率下降或者引起光抑制,净光合速率会出现下降的趋势。
植物的光饱和点(LSP)值是反映植物利用强光能力的一个重要指标,越低表明植物在强光条件下越易产生抑制效应,而光补偿点(LCP)较低则表明植物对弱光利用能力较强。刺槐的LCP值为1.9 μmol·m-2·s-1,LSP为1 499.2 μmol·m-2·s-1。
2.6 光-水分利用效率响应曲线
刺槐的光-水分利用效率响应曲线与光响应曲线的变化趋势很相似(图3),当PAR大于光补偿点时,随着PAR的增加,水分利用效率(WUE)快速增加,当光合有效辐射(PAR)接近光饱和点时,水分利用效率(WUE)达最大值,并基本呈较稳定的状态,随后光合有效辐射(PAR)继续增加,水分利用效率(WUE)反而下降。
2.7 固碳释氧量和降温增湿量及能力评价
2.7.1 日固碳释氧量和降温增湿量
利用采集的数据及相关公式计算得到刺槐植物8月份叶片的固碳(CO2)量和释氧(O2)量分别为2.93 g·m-2·d-1,1.77 g·m-2·d-1;蒸腾速率为1.16 mmol·m-2·s-1,释水量为 902.02 g·m-2·d-1,吸热量为2 254.54 kJ·m-2·d-1。
2.7.2 固碳释氧和降温增湿能力评价
利用单位叶面积年固碳(CO2)量和释氧(O2)量、年蒸腾水量和蒸腾吸热量来对刺槐的固碳释氧能力和降温增湿进行评价。固碳释氧能力:> 3 500 g为强、2 500—3 500 g为较强、<2500g 为弱;降温增湿能力:>350 kg为强、250—350 kg为较强、< 250 kg 为弱。若一年按 365 d计,则年固碳量和年蒸腾水量的计算公式如下:
年固碳量=日平均固碳量×365(d)
(10)
年蒸腾水量=日平均蒸腾水量×365(d)
(11)
计算得刺槐的年固碳量为1 069.45g,年蒸腾水量为 329.24 kg。因此,刺槐的固碳释氧能力较弱、降温增湿效益较强。
3 结论与讨论
本研究中,刺槐的净光合速率(A)日变化呈“双峰”曲线,第一个峰值出现在11:00为5.03 μmol·m-2·s-1,15:00出现第二个峰值,为2.22 μmol·m-2·s-1,具有明显的“午休”现象。Farquhar 等[16]认为,气孔限制和非气孔限制为影响A下降的主要因素。气孔限制为光照太强导致温度过高或过度失水,使得Gs减少甚至关闭,胞间 CO2浓度降低,影响了CO2的进入,使A下降;非气孔限制是因为叶肉细胞光合能力的下降,使叶肉细胞利用CO2的能力降低,最终胞间 CO2含量升高,A下降。结合刺槐的众多外部环境生态因子和内部生理因子日变化趋势和相关性系数,研究发现A与Gs呈显著正相关,表明影响刺槐的净光合速率的主要原因为气孔因素,这也说明其叶片光合“午休”是由气孔关闭或部分关闭引起的,与谢心等[17]的研究相同。
光补偿点较低、光饱和点较高的植物能够适应更宽的光照环境,而补偿点较高、光饱和点较低的植物对光照的适应性较窄[18]。刺槐的光补偿点低、光饱和点高,表明它们适应多种环境中的光照条件,对光的适应范围较宽,应属于喜阳类型,这与Bolat等[19]的研究一致。王庆红[20]在坡向和土壤质地对刺槐生长量的影响研究中认为:阳坡、半阴坡向和土壤质地有利于刺槐生长量的提高,而阴坡虽然土壤含水量较高,但生长量最低。因此,刺槐不适合在阴面生长,在种植人工林时可以将喜阴和喜阳植物结合,提高生态效应。
高净光合速率(A)和高水分利用率(WUE)可作为植物生存和广泛分布能力强弱的指标[21]。A反应出植物的光合能力的强弱,即在相同条件下,A值高的植物具有更强的光合能力和对环境的适应能力[22];而在相同条件下,水分利用效率(WUE)高的植物的抗旱能力较强。研究表明,刺槐的WUE随PAR的变化曲线与A随PAR的变化曲线相似,在相同条件下,A高的同时,WUE也高。因此,刺槐的抗旱能力和对环境适应能力较强,非常适合作为黄土高原地区防止水土流失的主要树种。
光合作用能利用光能把 CO2和 H2O合成糖及淀粉等有机物,光合作用产生的有机物质中有90%直接或者间接成为植物体内的干物质[23],而10%来自根部吸收的无机物质。因此,具有较高净光合速率的植物,其固碳释氧的生态效益相应的也较好。本研究通过对刺槐净光合速率日变化的测定计算了其固碳释氧的效益,发现刺槐的固碳释氧能力较弱。王丽勉等[24]研究了上海地区151种绿化植物的固碳释氧能力,并依据植物单位叶面积日吸收CO2释放O2量进行了归类,其中刺槐固碳释氧能力中等。这些结论与本研究结果存在一定的差异,可能与延安特有的土质以及环境有关。
光合作用和蒸腾作用吸收大部分照射到绿色植物叶面上的太阳辐射,而被吸收的太阳辐射可以大部分转化为植物蒸腾作用吸收的热量。根据植物的这个特点,可以通过测定植物的蒸腾速率来评价其降温增湿能力。本项目测定8月份刺槐的年蒸腾水量为 329.24 kg(250—350 kg),刺槐的降温增湿效益较强。但与深圳市的28种植物[10]相比,仍处于较低水平,这跟延安地区土壤含水量较低以及刺槐自身的生理调控力有一定的相关性。
刺槐的优点为对光的适应范围较宽、抗旱能力和对环境适应能力较强、降温增湿能力较强,缺点为其净光合速率(A)受气孔因素较大,固碳释氧能力较弱。因此,在人工林的选择和配置过程中,不能只单一的种植刺槐,可以通过配置复合结构,增加单位叶面积指数,增强单位面积的生态效应,从而提高整个绿地系统的生态效应。
猜你喜欢
节水灌溉(2021年5期)2021-05-26 06:49:38
现代园艺(2020年2期)2020-12-22 05:57:04
中国农业科技导报(2020年12期)2020-03-15 10:50:08
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
现代园艺(2018年2期)2018-03-15 08:00:12
新课程·下旬(2017年7期)2017-08-14 11:12:50
山东林业科技(2016年5期)2016-07-05 00:43:00
西北林学院学报(2015年5期)2015-01-03 05:12:20
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
植物营养与肥料学报(2011年2期)2011-10-26 03:52:46
