
树鼩IgE高亲和力受体α亚基的原核表达、抗体制备及鉴定
2018-10-23 11:27张晶晶肖红剑李育中宫悦毕研伟寸韡
生物技术通讯 2018年5期
张晶晶,肖红剑,李育中,宫悦,毕研伟,寸韡
中国医学科学院 北京协和医学院 医学生物学研究所,云南省重大传染病疫苗研发重点实验室,云南昆明650118
随着卫生条件的改善,花粉过敏、哮喘等过敏性疾病患者在发达国家逐年增加,甚至有研究者提出了“卫生假说”,认为卫生条件越好儿童早期感染越少,后期患过敏的几率越大。“卫生假说”目前仍存在争议,而过敏性疾病的发病机理还有待深入研究。通常速发型过敏症状是由Ⅰ型超敏反应引起的,过敏原第一次侵入机体可以诱导B淋巴细胞产生抗原特异性IgE,IgE与靶细胞表面的高亲和力受体(FcεRI)结合后,当相同抗原再次侵入机体时,就可以引发靶细胞内的一系列信号转导,使靶细胞释放组胺等生物活性物质,引起过敏性炎症反应。FcεRI是四聚体结构,由胞外α链、β链、γ-γ链组成[1],在过敏反应中与IgE结合的是其α亚基(FcεRIα)[2]。目前,研究Ⅰ型过敏反应的小动物模型主要是啮齿类动物,但是由于物种差异较大,很多研究结果与人体存在差异。近年来,树鼩作为一种新型实验动物正日益受到关注。作为灵长类动物的近亲,树鼩在生理解剖、神经发育、病毒感染特性及心理应激模式等方面与灵长类甚至人类高度相似[3]。树鼩全基因组测序分析结果显示,其免疫系统与灵长类动物的基因组具有较高的相似性[4-5];同时,树鼩的高亲和力受体也存在α亚基,但对树鼩FcεRIα的相关研究较少。为了更好地建立研究树鼩过敏反应的动物模型,我们调取了树鼩FcεRIα基因并表达该受体蛋白,免疫动物制备抗体,为树鼩过敏动物模型的建立奠定了基础。
1 材料与方法
1.1 材料
SPF级2~3周龄雄性树鼩,体质量130~180 g,购于中国医学科学院医学生物学研究所国家树鼩种质资源中心;清洁级雌性新西兰大白兔2~2.5 kg,购于昆明医科大学实验动物中心;大肠杆菌DH5α、BL21,质粒载体pET-30a(+)由本室保存。
限制性内切酶HindⅢ、NdeⅠ、XhoⅠ,T4DNA连接酶等购自NEB公司;TRIzol裂解液、RT-PCR反转录试剂盒购自上海生工公司;TaqDNA聚合酶购自南京诺唯赞生物科技公司;质粒大提试剂盒、质粒小提试剂盒及通用性DNA纯化回收试剂盒购自北京天根技术公司;弗氏完全佐剂、弗氏不完全佐剂购自Sigama公司;HPR标记的羊抗兔IgG、Cy3标记的驴抗兔IgG购自Proteintech公司;转染试剂jetPrime购自Polyplus公司。
1.2 树鼩FcεR1α基因的调取
通过NCBI调取树鼩相关基因(GenBank:XM_014584294.1)(1008 bp)为预测参考序列,设计引物进行巢式PCR,验证基因序列是否正确。Nuno⁃mura[6]等曾在小鼠肝脏中提取过IgE受体基因,因此本研究用TRIzol法快速提取RNA试剂盒提取树鼩的脾脏和肝脏RNA,逆转录成cDNA进行巢式PCR。巢式引物设计如下:
Primer1-F:5′-TCTCTCCAGACCAGTCAGTACTTATTC-3′;Primer1-R:5′-TCCTGAGCACATATTTTTCTATGTACG-3′;Primer2-F:5′-CCAGTTTTGAACGCAGAGATCTC-3′;
Primer2-R:5′-GCAGTTGCTGATGCTAGAAATCGC-3′。
1.3 树鼩IgE FcεR1α序列信号肽及跨膜区的预测及表达质粒构建
用SignalP 4.1信号肽预测软件和TMHMM跨膜预测软件预测人类和树鼩的IgE FcεRIα的信号肽和跨膜区序列。以pET-30a(+)为载体设计目的基因序列的引物,构建pET-30a(+)/ts⁃FcεR1α和 pAAV-CMV-tsFcεR1α质量。引物为Primer3-F(5′-GAACATATGGCCCTCCAGAAAACG GTC-3′;GAA为保护碱基,划线部位为NdeI酶切位点)、Primer3-R(5′-GCAAAGCTTGGTATTCTTG CTGTGCTGAGC-3′;GCA为保护碱基,划线部位为HindⅢ酶切位点)。
1.4 树鼩IgE FcεRIα的诱导表达
将构建正确的质粒转化大肠杆菌BL21(DE3),涂板培养,挑取单克隆,37℃振荡培养至菌液D600nm值为0.7时,用IPTG(终浓度为1 mmol/L)诱导表达,每隔1 h取样,菌液超声波破碎,通过SDS-PAGE找出最佳诱导时间。将诱导4 h的菌液离心,超声波破碎,4℃、8000 r/min离心20 min,SDS-PAGE显示蛋白主要表达在沉淀中,然后扩大培养制备包涵体。
1.5 树鼩IgE FcεRIα的纯化
称取包涵体,用6 mol/L盐酸胍在37℃水浴中溶解包涵体约30 min,4℃、8000 r/min离心10 min,将溶解的包涵体样品在冰上稀释复性,并过夜放置复性,复性液用超滤器浓缩超滤,浓缩后的复性液通过AKTA explorer蛋白纯化系统进行纯化(平衡柱子→上复性液→结合液平衡柱子→洗脱液洗脱目的蛋白),将洗脱的目的蛋白进行SDS-PAGE检测。
1.6 树鼩IgE FcεRIα多克隆抗体的制备
将纯化后的树鼩IgE FcεRIα与等体积的完全弗氏佐剂充分乳化混合后,于背部皮下多点第1次免疫新西兰大白兔,所用蛋白量为400 μg;之后每隔2周分别进行第2次和第3次免疫,将等体积的蛋白和不完全弗氏佐剂充分乳化混合后进行免疫;第3次免疫后颈动脉采血,分离血清。
1.7 树鼩IgE FcεRIα多克隆抗体的特异性及效价检测
1.7.1 Western印迹检测 为了验证多克隆抗体的特异性,用制备的多克隆抗体鉴定pET-30a(+)/tsFcεR1α重组质粒诱导后的菌液,进行West⁃ern印迹检测。同时为了验证多克隆抗体是否能在真核细胞检测到表达的树鼩FcεR1α,用质粒pAAV-CMV-tsFcεRIα转染293细胞,通过Western印迹检测制备的多克隆抗体的生物学活性。
1.7.2 细胞免疫荧光检测 293细胞接种至24孔板,待细胞密度达到70% 左右,用质粒pCW711(pAAV-CMV-tsFcεRIα)转染细胞,48 h后将细胞上清吸出,用4% 多聚甲醛固定10 min,PBS洗涤;1% BSA室温封闭30 min,PBS洗涤;孵育一抗,按1∶2000室温孵育30 min,PBS洗涤;孵育荧光二抗(Cy3标记的驴抗兔IgG),按1∶2000室温避光孵育30 min,PBS洗涤;滴加DAPI复染核并进行封片,通过荧光显微镜观察。
1.7.3 ELISA检测多克隆抗体效价 将纯化的蛋白稀释至 2 μg/mL,每孔包被 100 μL,4℃过夜,1% BSA室温封闭1 h,PBST洗涤1次,将兔血清分别稀释至1/100、1/10 000、1/20 000、1/50 000、1/100 000、1/200 000、1/500 000,每孔孵育 100 μL,37℃孵育1 h,PBST洗涤;孵育HRP标记的羊抗兔(1∶5000)100 μL/孔,37℃孵育40 min;PBST洗板3次,滴加100 μL显色液(TMB终浓度0.12 mg/mL,过氧化氢终浓度0.009% ),室温避光显色5~10 min,加入50 μL终止液(2 mol/L硫酸)终止反应,于450 nm检测吸收峰。
2 结果
2.1 克隆树鼩IgE FcεRIα cDNA片段
以树鼩的脾脏和肝脏为实验材料提取RNA,反转录合成第一链cDNA,巢式PCR(扩增条件:95℃ 30 s预变性;95℃ 10 s变性、56℃ 30 s退火、72℃ 30 s延伸,35个反应循环;最后72℃延伸2 min)结果表明该基因在树鼩脾脏中含量较高,因此后续试验采用树鼩脾脏部位进行。将PCR产物连接T载体进行测序,测序结果与树鼩IgE FcεRIα序列比对基本一致。结果见图1。
2.2 树鼩IgE FcεR1α的信号肽及跨膜区预测
NCBI公布的数据表明人类IgE FcεRIα的信号肽序列为1~25氨基酸残基,跨膜结构域为203~225氨基酸残基。用SignalP 4.1软件预测的人类IgE FcεRIα信号肽序列也为1~25氨基酸残基,用TMHMM跨膜软件预测的跨膜结构域为203~225氨基酸残基;同时用SignalP 4.1和TMHMM软件预测树鼩IgE FcεRIα的信号肽及跨膜结构域序列,信号肽为1~25氨基酸残基,胞外结构域为1~213氨基酸残基,跨膜结构域为214~236氨基酸残基。因此,本实验扩增的序列为胞外结构域去信号肽的第26~213氨基酸残基,预测结果见图2。
2.3 构建重组表达质粒
用设计的引物扩增编码26~213氨基酸残基的基因序列,目标片段存在非特异性扩增,经切胶纯化回收,用NdeⅠ、HindⅢ对目的片段和pET-30a(+)载体双酶切,T4DNA连接酶16℃连接过夜,将构建的质粒用NdeⅠ、HindⅢ酶切,获得569和5621 bp的片段,表明构建了原核重组表达质粒 pET-30a(+)/tsFcεRIα。 将 pET-30a(+)/ts⁃FcεRIα及载体 pAAV-CMV 用EcoRⅠ和SalⅠ酶切,T4DNA连接酶16℃连接过夜,将构建的质粒用BamHⅠ酶切,获得1045和5026 bp片段,表明真核表达质粒构建成功。结果见图3。
2.4 树鼩IgE FcεRIα重组蛋白的表达及纯化
将构建的质粒测序,测序结果与树鼩该基因比对序列一致;构建的质粒转化大肠杆菌BL21,涂板,37℃振荡培养,菌液D600nm为0.7时用IPTG诱导表达,分别在1、2、3、4、5 h取样,菌液超声波破碎后进行SDS-PAGE,结果显示在诱导4 h后蛋白表达量没有明显增加,因此确定第4 h为蛋白最佳诱导时间。将制备好的包涵体用6 mol/L盐酸胍在37℃水浴中溶解约30 min,4℃、8000 r/min离心10 min,去掉部分沉淀,将溶解的包涵体样品在冰上边滴加边搅拌稀释复性,复性液为20 mmol/L Tris-HCl(pH8.8)、1% 精氨酸、1 mmol/L还原型谷胱甘肽,过夜放置复性,将复性液过已经处理好的Ni柱进行亲和层析纯化,洗脱液经SDS-PAGE,蛋白纯化率约90% 。结果见图4。
2.5 树鼩IgE FcεRIα多克隆抗体的制备及检测
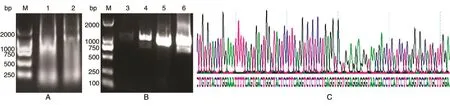
图1 树鼩总RNA提取及PCR扩增
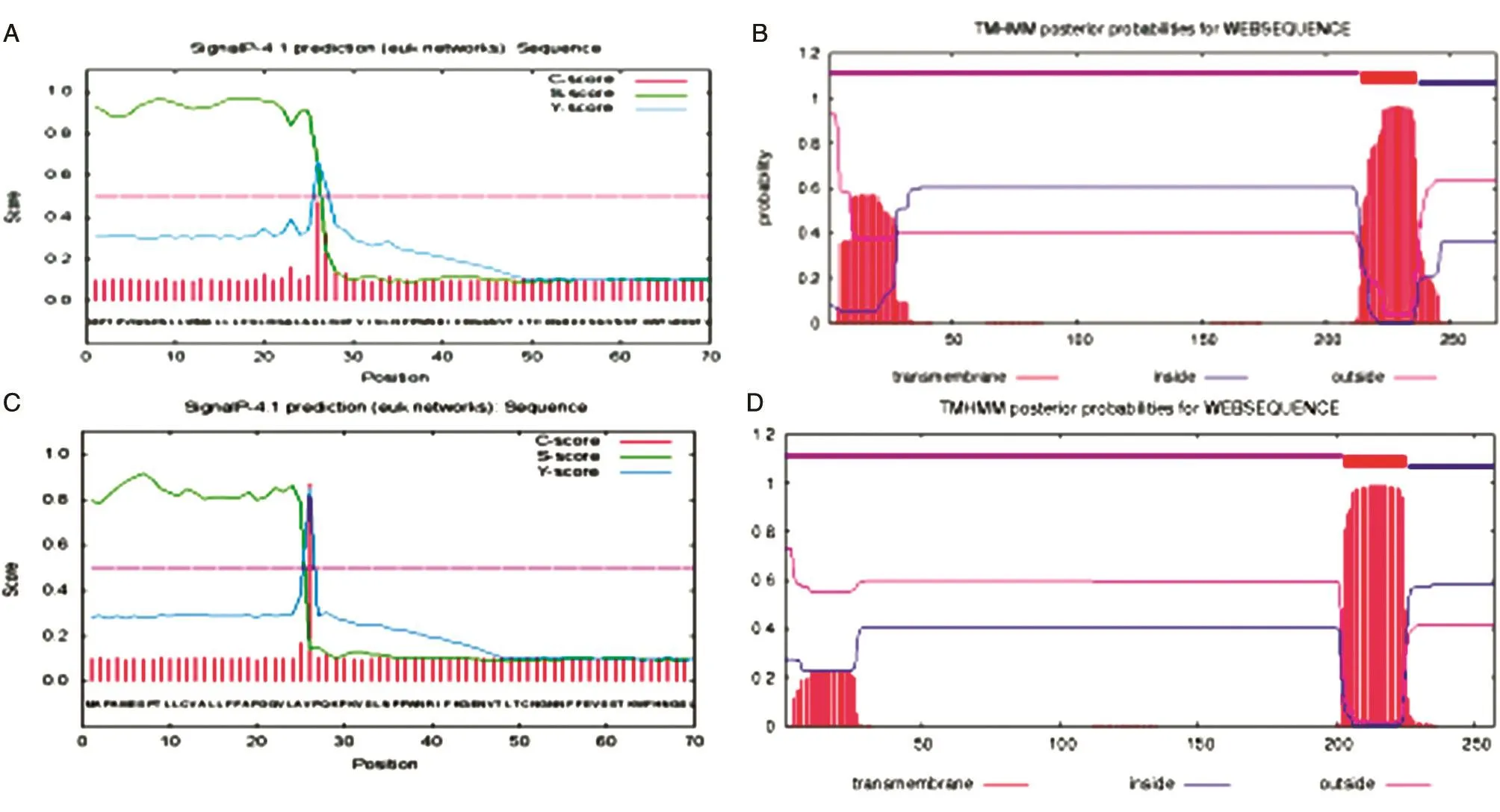
图2 树鼩和人IgE FCεR1α的信号肽及跨膜区预测
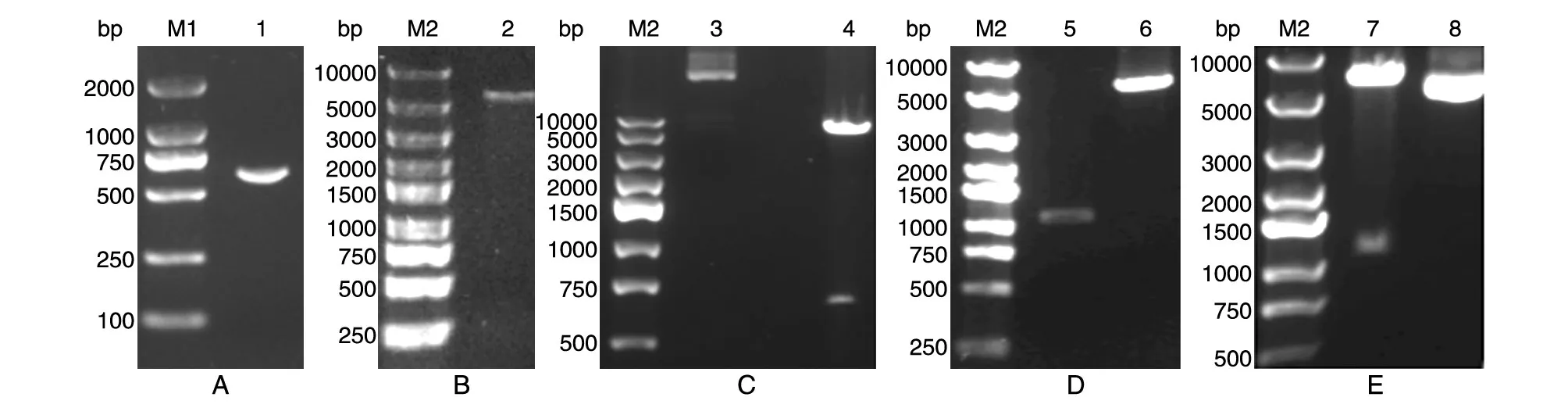
图3 重组表达质粒的构建及鉴定
Western印迹(图5A)表明,未经IPTG诱导的菌液中未检测到原核表达的IgE FcεR1α,而IPTG诱导后的菌液中能检测到且条带单一,证明制备的抗体特异性较好。在真核系统中表达的蛋白存在翻译后修饰,IgE FcεR1α比原核系统表达的偏大,且该抗体能检测到真核细胞中表达的IgE FcεR1α,条带单一,特异性较好(图5B)。免疫荧光结果证明真核细胞中表达的该蛋白位于细胞膜表面,阴性对照未检测到(图5C)。ELISA检测该抗体的效价可达到100 000(图5D)。
3 讨论
过敏性疾病是常见疾病,临床症状多样,包括过敏性哮喘、过敏性休克、过敏性鼻炎、过敏性皮炎等。在过去的20年中过敏性疾病的发病率逐年上升,全世界有1.0~1.5亿人患有哮喘,每年可导致约18万人死亡。过敏性疾病是由Ⅰ型超敏反应引起的,其主要中间物质为IgE及其高亲和力受体FcεRI,FcεRI主要分布于肥大细胞和嗜碱性粒细胞中。IgE与FcεRIα具有较高的亲和力(亲和常数Ka≈10-10mol/L),二者结合可引起下游级联反应,从而引发炎症反应[7]。在小鼠哮喘模型中,IgE缺陷型或FcεRIα受体缺陷型接受尘螨提取物均未表现严重的气道高炎症反应,相反,IgE和FcεRIα均独立负责气道高炎症反应[8-9]。因此,FcεRIα是参与Ⅰ型超敏反应的关键分子。随着分子免疫技术的发展,对抗体-受体相互作用的分子机制有了深入的了解,可以通过人工合成针对FcεRI特异性高、亲和力强,但与其作用不产生生物学活性的抗体片段,从而对体内的IgE产生竞争性抑制作用,达到治疗目的[10-12]。李莉等[13]已制备出膜结合型抗大鼠FcεRIα的单克隆抗体,并证实其能竞争性抑制IgE与肥大细胞的结合。利用非过敏原性的抗IgE高亲和力单克隆抗体与体内游离的IgE结合形成的免疫复合物能够中和IgE,抑制IgE合成,从而降低肥大细胞和嗜碱性粒细胞上IgE受体的表达[14-16]。IgE FcεRIα抗体还可作为检测及筛选工具,通过流式细胞仪筛选肥大细胞和嗜碱性粒细胞等。目前,用于过敏反应的动物模型基本局限于小鼠,通过全基因组序列进行生物进化树分析,发现树鼩比小鼠更接近人类,树鼩与人类在基因方面具有遗传关系[17]。目前,关于树鼩的过敏反应机制研究尚少。我们表达了树鼩IgE FcεRIα蛋白,并制备出多克隆抗体,为今后研究树鼩高亲和力受体及建立树鼩过敏反应动物模型奠定了基础。
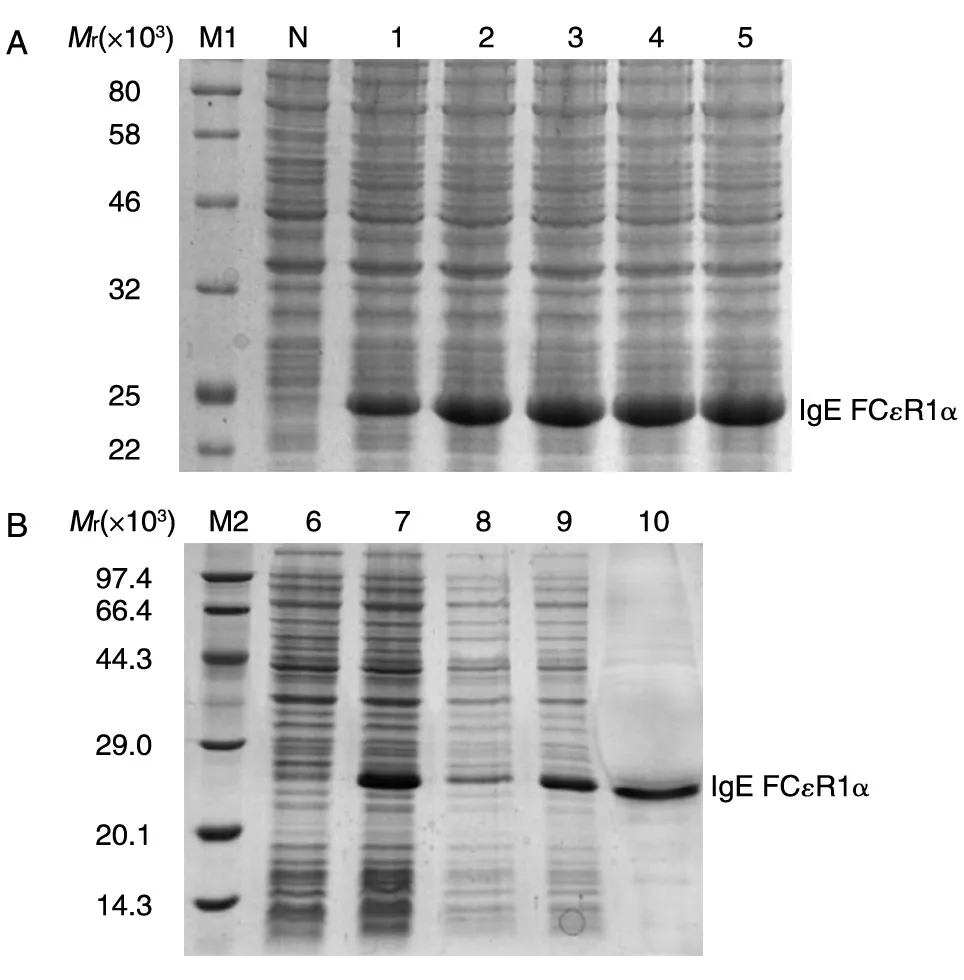
图4 FCεR1α诱导表达与纯化的SDS-PAGE
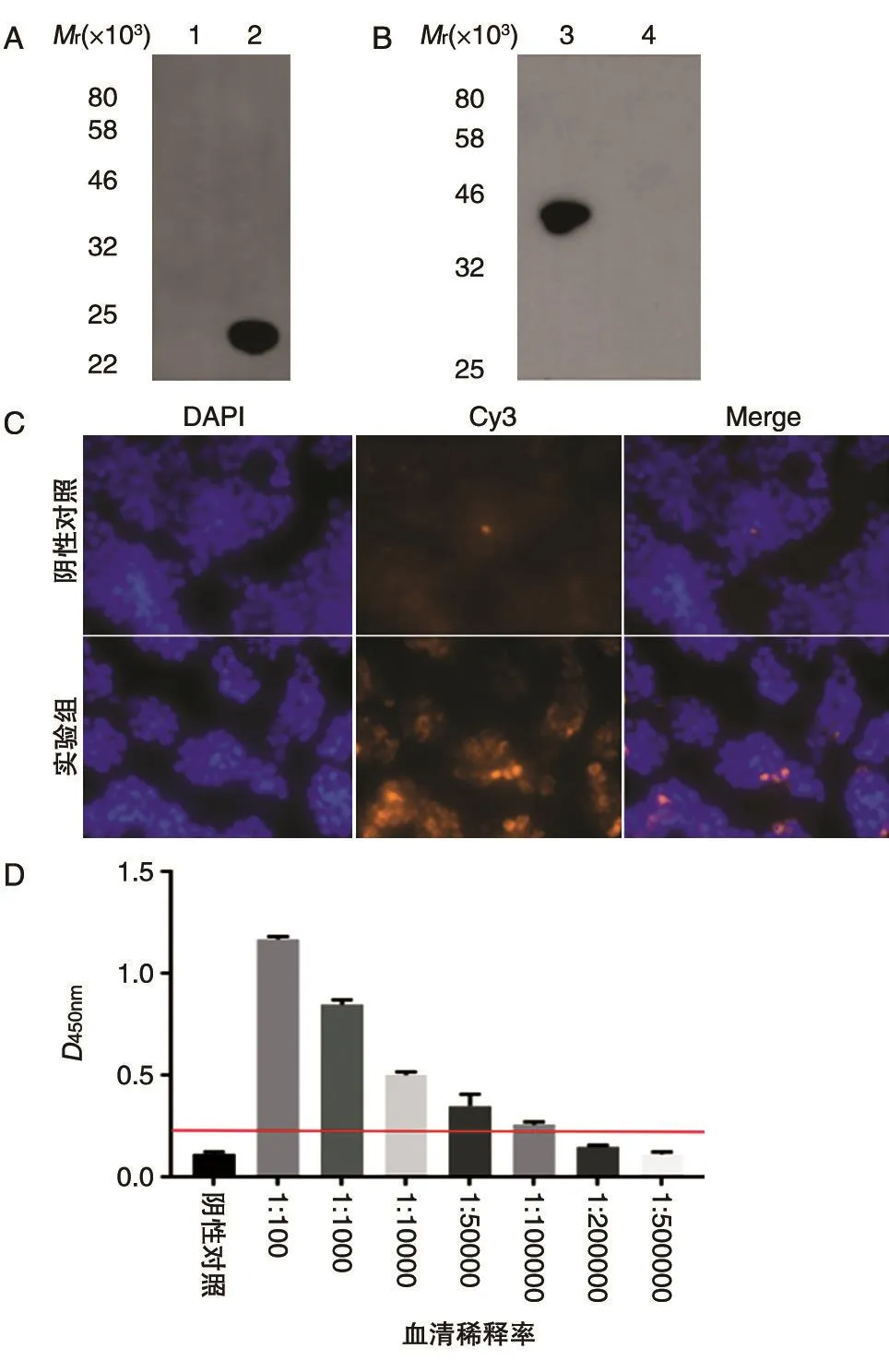
图5 树鼩IgE FCεR1α多克隆抗体的检测
猜你喜欢
生物化学与生物物理进展(2022年6期)2022-07-21
生物技术通报(2021年6期)2021-08-11
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
商品与质量(2018年38期)2018-04-16
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
池州学院学报(2015年3期)2016-01-05
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
天津科技大学学报(2015年2期)2015-08-09
大连工业大学学报(2014年4期)2014-09-19
