
胃蝇属三龄幼虫形态特征及行为的种间和种内关系分析
2018-10-15 10:26:50刘善辉胡德夫范晓泽
中国兽医杂志 2018年6期
刘善辉,胡德夫,李 凯,范晓泽
(1.北京语言大学教务处,北京海淀100083 ; 2.新疆农业大学动物医学学院,新疆乌鲁木齐8300523 ;3.北京林业大学自然保护区学院,北京海淀100083 ; 4.中国动物园协会,北京西城100037)
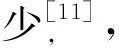
1 材料与方法
1.1 材料来源 马胃蝇L3来源有三:其一,患胃蝇蛆病的野马驱虫所得,所用药品为“易杀得”,由北京万丰药业有限公司生产,有效成分伊维菌素;其二,剖检当地家马消化道所得;其三,马科动物自然排出粪便中所得。
1.2 测量方法 所得幼虫均参考文献记载[2]使用光学显微镜(莱卡MZ75)观察。采用游标卡尺(精度0.01 mm)和电子天平(精度0.001 g)测量和称重L3体长、体宽和体重。使用带有刻度的大号透明玻璃容器进行L3的入土行为观察,以幼虫体表完全没入土表层为基准记录入土时间,以入土24 h时间为间隔(认为幼虫不继续入土),测量胃蝇L3的入土深度。
1.3 统计分析 数据处理采用SPSS22.0软件对L3的表型性状进行方差分析(ANOVA)、变异系数(CV) 差异性检验、多重比较分析(Tukey-HSDa)、相关分析(Person-Correlation)。并依据表型数据和形态特征矩阵及行为特点对L3按非加权配对算术平均法(Un-weighted pair-group method using arithmetic averages,UPGMA)进行系统聚类分析。
2 结果与分析
2.1 马胃蝇L3基本形态特征 以马胃蝇体色、棘刺形状、棘刺有无、口钩形状等特征建立胃蝇属三龄幼虫表观矩阵,具体如表1所示。
2.2 马胃蝇L3形态数量性状的种间和种内变异 经方差分析可知,L3种间体长(F = 33.084,P<0.05)、体宽(F = 130.921,P< 0.05)、体长/体宽(F = 72.388,P<0.05)、体重(F=88.073,P<0.05)差异均达到显著水平。多重比较结果(表2)显示,马胃蝇L3体长除在黑腹胃蝇和红尾胃蝇、红尾胃蝇和黑角胃蝇,鼻胃蝇和裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05); L3体宽除在肠胃蝇和黑角胃蝇、黑腹胃蝇及红尾胃蝇和裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05);L3体重除在肠胃蝇和黑角胃蝇及鼻胃蝇、黑腹胃蝇及红尾胃蝇和裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05);L3体长/体宽除在红尾胃蝇和黑腹胃蝇及肠胃蝇和裸节胃蝇、裸节胃蝇及鼻胃蝇和黑角胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05)。L3种间表征总体变异系数为体重(22.75%)> 体长/体宽(19.16%)>体宽(13.95%)>体长(13.03%)。各个表征的变异系数存在差异,其中肠胃蝇种内L3体长、体宽、体重及体长/体宽的变异系数均小于总体变异系数;黑腹胃蝇和鼻胃蝇的体长、体宽、体重及体长/体宽的变异系数均大于总体变异系数;裸节胃蝇体长、体宽及体长/体宽的变异系数均小于总体变异系数;红尾胃蝇的体宽、体重、体长/体宽的变异系数均小于总体变异系数。
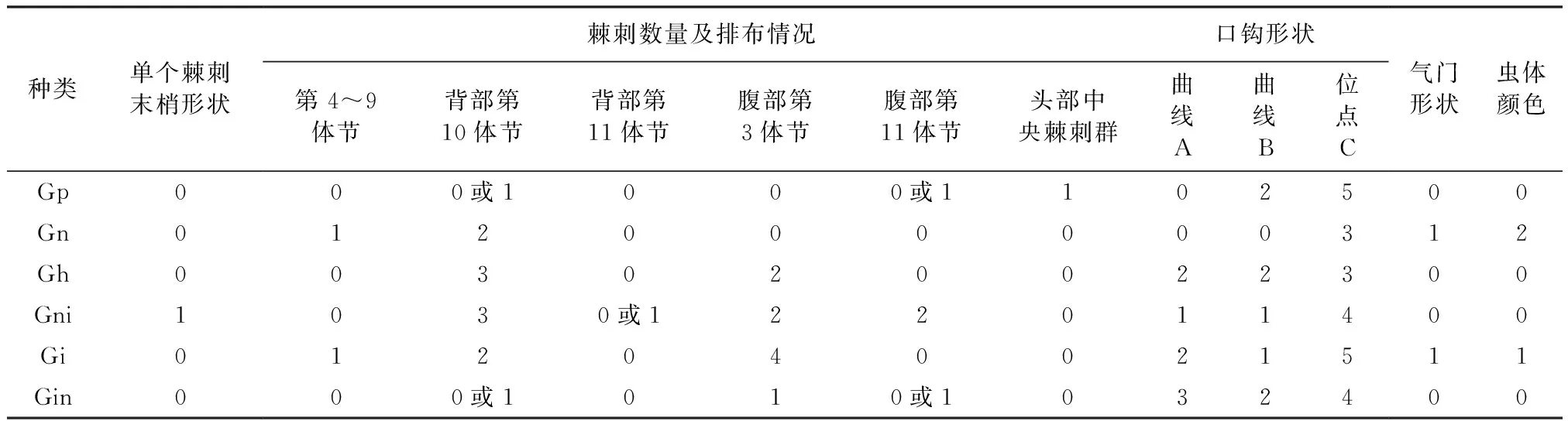
表1 马胃蝇表观性征的0~1矩阵
注:单个棘刺末梢形状(0=尖锐;1=钝);第4至9体节棘刺排数(0=双排;1=单排);背部第10体节棘刺排布(0=无;1=两侧少许;2=单列中间断开;3=双列中间断开);背部第11体节棘刺排布(0=无;1=两侧少许);腹部第3体节棘刺排布(0=单列中间不断开;1=单列中间断开;2=双列中间不断开;3=双列中间断开;4=无);腹部第11体节棘刺排布(0=无;1=单列;2=双列);头部中央棘刺群排布(0=缺失;1=存在);口钩形状(0=轻微凹陷;1=强烈凹陷;2=平直; 3=轻微突出;4=强烈突出;5=向内弯曲);后气门形状(0=耳形;1=肾形);虫体颜色(0=红色;1=绿色;2=黄色)。黑腹胃蝇(Gp,Gasterophiluspecorum)、肠胃蝇(Gi,G.intestinalis)、裸节胃蝇(Gin,G.inermis)、黑角胃蝇(Gni,G.nigricornis)、红尾胃蝇(Gh,G.haemorrhoidalis)、鼻胃蝇(Gn,G.nasalis)
2.3 马胃蝇L3行为性状的种间和种内变异 方差分析结果(表3)显示,马胃蝇L3种间爬行蠕动频率(F = 82.620,P<0.05)、爬行速度(F = 63.296,P<0.05)、入土深度(F = 7.111,P<0.05)、入土时间(F = 30.814,P<0.05)差异均达显著水平。多重比较结果显示,L3蠕动频率除在黑角胃蝇和鼻胃蝇、黑腹胃蝇和肠胃蝇,肠胃蝇和红尾胃蝇及裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05);爬行速度除在黑腹胃蝇和黑角胃蝇、红尾胃蝇和肠胃蝇及裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05);入土深度除在黑腹胃蝇和红尾胃蝇及裸节胃蝇、红尾胃蝇和鼻胃蝇及裸节胃蝇、鼻胃蝇和肠胃蝇及裸节胃蝇、黑角胃蝇和肠胃蝇及裸节胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05);入土时间除在裸节胃蝇和红尾胃蝇及肠胃蝇间差异不显著外,在其余种间差异均达显著水平(P<0.05)。
表3显示,L3蠕动频率、爬行速度、入土深度和入土时间平均变异系数依次为:入土深度(25.25%)> 爬行速度(25.09%)> 蠕动频率(24.60%)入土时间(20.75%);蠕动频率变异系数则以鼻胃蝇和裸节胃蝇为最大和最小,分别为28.70%和14.14%,其对应变幅分别为8.0~22.0 s/次和3.0~4.0 s/次;爬行速度变异系数以肠胃蝇和黑角胃蝇为最大和最小,分别为36.00%和12.58%,其对应变幅分别为0.8~2.1 cm/min和3.4~5.6 cm/min;入土深度变异系数以鼻胃蝇和黑角胃蝇为最大和最小,分别为31.89%和20.08%,其对应变幅分别为1.2~4.2 cm和1.1~2.4 cm;入土时间变异系数以黑腹胃蝇和裸节胃蝇为最大和最小,分别为30.08%和14.75%,其对应变幅分别为1.2~5.0 min和5.2~7.5 min。
2.4 马胃蝇L3数量性状和行为性状间的相关性分析 对马胃蝇数量性状和行为性状进行相关分析,结果显示:马胃蝇种间L3体长与爬行速度和入土深度分别呈显著负相关(R=- 0.126,P< 0.05)和极显著负相关(R=- 0.171,P< 0.01);L3体宽与蠕动频率呈显著正相关(R=0.133,P<0.05);L3体重与爬行速度及入土深度分别呈极显著负相关(R=- 0.179,P<0.01)和显著负相关(R=- 0.138,P<0.05);L3体长/体宽与入土时间呈显著正相关(R=0.141,P<0.05),与蠕动频率和爬行速度分别呈极显著负相关(R=- 0.197,P<0.01)和显著负相关(R=- 0.138,P<0.05)。
2.5 马胃蝇L3的聚类分析 根据其12个形态形状和4个数量形状及4个行为学形状对6个马胃蝇种L3进行聚类分析,如图1所示:以欧氏距离10为阀值,胃蝇L3划分为3簇,其中红尾胃蝇、肠胃蝇、裸节胃蝇为一簇,鼻胃蝇和黑角胃蝇为一簇,黑腹胃蝇为一簇。
3 讨论与结论
变异是生物界所具有的最基本特征之一。在诸多变异之中,表型性状变异既具有变异性又具有稳定性,对表型性状变异的描述和分析是探讨其他变异类型的基础。马胃蝇L3作为胃蝇生活史繁衍过程中一个重要的过渡虫态,其形态的差异反映了对环境的适应,目前对其L3表征变异模式的研究较少。马胃蝇L3表征是环境和遗传因子相互作用的结果,既表现出对环境的适应与响应特征,又表现出其固有的形态特征。如马胃蝇L3在颜色、棘刺大小、口钩形状、棘刺先端尖或钝等表征受种间强的遗传控制,表现出固有的形态特征,是长期适应自然的结果。本研究发现胃蝇种间L3在体长、体宽、体重、体长/体宽和蠕动频率、爬行速度、入土深度及入土时间上均达到显著差异性水平,且体长(13.03%)和入土时间(20.75%)的变异系数最小,也是众多表征中相对最为稳定。
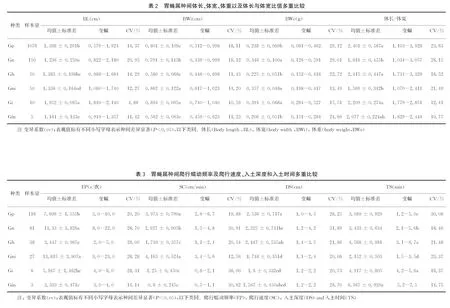
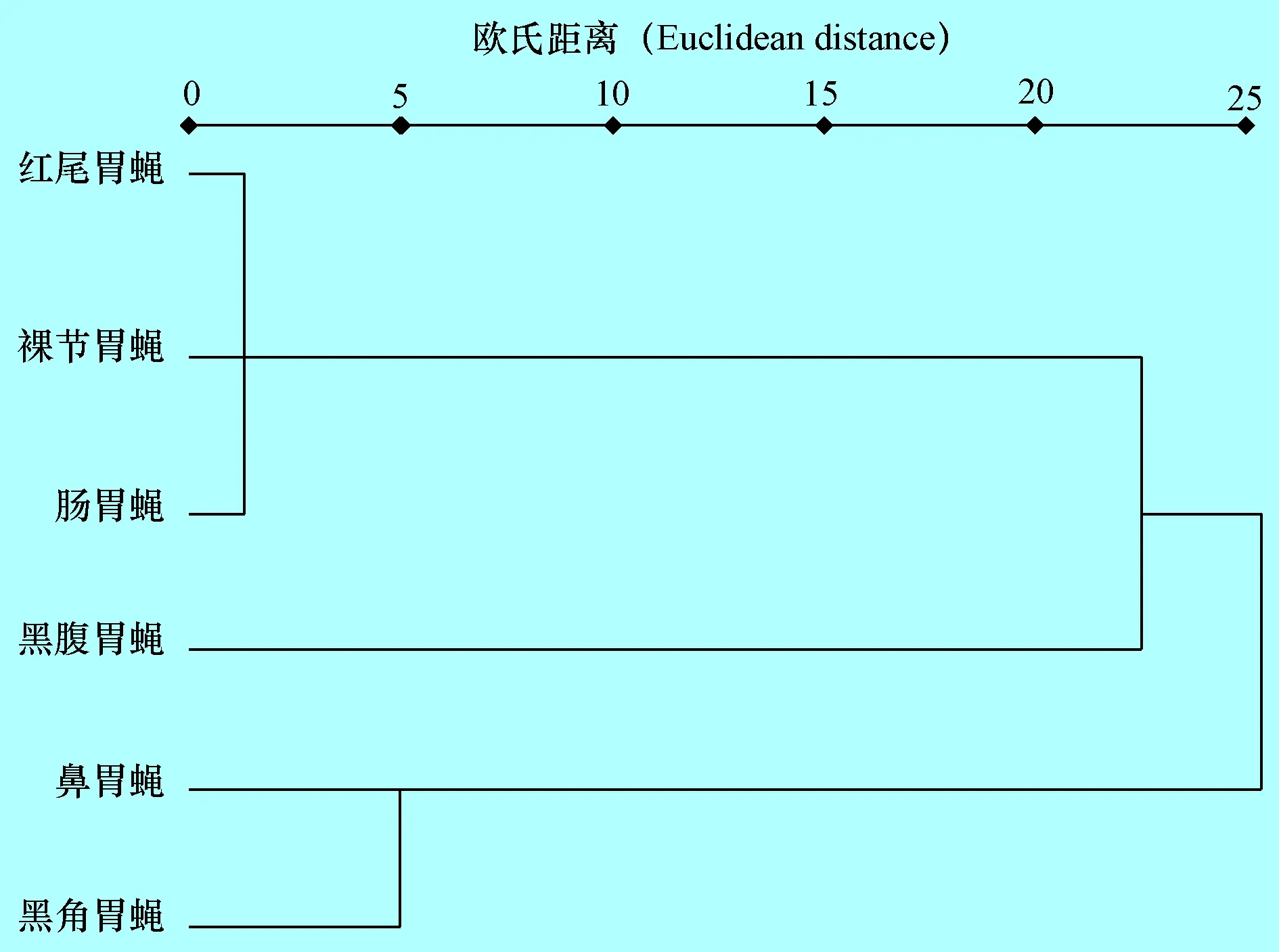
图1马胃蝇L3聚类分析
Valentine (1976)[12]指出,专食性物种可能存在较低的遗传变异,而杂食性的物种遗传多样性相对较高。然而,马胃蝇作为一种较为专一性的寄生虫,主要的幼虫期基本都在宿主体内度过,依靠宿主体内营养维持生命体征,面对宿主体内和体外复杂多变的生境中长期的适应,自身遗传力与外界因子的相互作用,却逐渐在种群内产生了较高水平的表型性状多样性。本研究发现马胃蝇L3的4个表型性状和4个行为性状间差异性显著,变异最大的马胃蝇种群是黑腹胃蝇 ( CV= 29.12%),说明黑腹胃蝇的种群遗传多样性较高。推测其表型性状遗传多样性高的原因应与其所处气候类型及其他生态因子有关。KNR是众多马科动物同域生活的区域,互相之间发生交叉感染几率较大,非常适宜马胃蝇的生长和发育。有研究表明,黑腹胃蝇成虫的平均寿命、产卵量和繁殖力都高于其他种马胃蝇[1]。
表型性状的变异受其自身的遗传组成和所处生态环境共同影响,气候因素会对昆虫的分布产生直接和间接的影响。对于寄生虫而言,多种环境因子[4]及宿主活动范围[13]可能会成为影响其分布的主要原因,尤其是温度和湿度[14]。本研究发现马胃蝇L3体长、体宽、体重及体长/体宽表征在同一环境条件下差异显著,且众多表征间存在显著正相关,推测不同L3形态特征除环境影响外主要受遗传控制,上述形态特征之间存在相互促进、共同变异的关系。变异系数的大小表明了表征的离散程度,变异系数越大,表示其值的离散程度越大,具有丰富的遗传变异,变异系数越小,表明表征较稳定。形态特征是与行为特性之间存在密切联系,其特定结构和特殊行为都是为了能够更好的适应周围环境。马胃蝇L3体长和体宽与攀附宿主体壁所占表面积及空间有关,体长与体宽的特定比值能够更加有利于其在有限的宿主体内获取生存空间;入土的深度及入土时间与L3化蛹期长短及被天敌发现几率有关,如果入土时间过长则暴露在外被天敌发现几率会不断增加,并且不能够为化蛹提供一个相对稳定的生境;蠕动频率及爬行速度都与其能否迅速入土,避开天敌捕食有关。
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
天天爱科学(2022年9期)2022-09-15 01:12:28
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
奇闻怪事(2019年8期)2019-08-30 04:57:39
科学之友(2019年1期)2019-02-24 08:22:24
作文评点报·中考版(2017年1期)2017-03-06 21:37:28
杂草学报(2015年2期)2016-01-04 14:57:55
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:02
