
干旱条件对鬼针草和醉鱼草种间相互作用及生长的影响
2023-09-09 08:01:22陈家兴陈林丽侯夏丽杨庆祝尹任娅
植物研究 2023年5期
陈家兴 王 姝 陈林丽 侯夏丽 杨庆祝 尹任娅
(贵州大学林学院,贵阳 550025)
植物种间相互作用广泛地发生于自然界[1],其相互作用的结果包括竞争和促进[2]。而非生物环境条件能改变相互作用的结果[3],如水分条件,在半干旱草原中不同植物,随着降雨量的不断减少,种间关系先由竞争转向促进,再变成竞争[4];或比较湿润和干旱环境的影响,湿润下植物竞争强于干旱环境,而干旱条件下植物的叶片能相互遮挡,并改善微气候从而减少叶片水分的流失,由此表现出促进作用[5]。然而衡量相互作用的指标往往是总生物量[6],对于植物根、茎、叶是否与总生物量响应一致,相关问题仍不清晰,如根的竞争强度可能强于总生物量[7]。此外,植物间相互作用的结果往往是竞争和促进作用的综合结果,非生物环境条件可能通过改变竞争与促进作用的相对强度从而影响植物不同构件特征。
水分是对植物存活、生长和繁殖起关键性作用的非生物环境因子之一,也会影响物种的分布格局[8-9]。水分能够改变植物生物量的分配[10],然而不同水分条件下受种间相互作用的影响,植物可能会采取不同生物量分配策略,如湿润条件下种间竞争降低了滨菊(Leucanthemum vulgare)和硫委陵菜(Potentilla recta)的根冠比,而干旱减弱根冠比对种间竞争的响应[11]。类似研究往往只关注根冠比的响应,而叶片、茎和根的结论基于根冠比的研究可能有很大不同[12]。另外,一些研究表明不同水分条件下,植物地上与地下部分对竞争的响应存在不一致的现象,说明植物种间作用会加剧或减弱根、茎和叶对干旱的响应[11]。考虑到植物种间作用对根、茎、叶的影响及与环境的关系,其结果可能阐明了物种共存过程中的适应策略。
关注外来植物的入侵能力,是入侵生态学研究的重点[13]。鬼针草(Bidens pilosa)是一种入侵性较强的植物,目前国内研究已从鬼针草环境适应性[14]、种子繁殖特性[15]、化感作用[16]等方面解释其入侵性,然而对于鬼针草相对生长率的研究较少。已有研究[17-18]表示入侵植物的相对生长速率也是其入侵性的重要表现。因此,研究干旱地区先锋植物鬼针草[19]的相对生长率,可以了解鬼针草的生长规律。另外,鬼针草作为一种广泛入侵的恶性杂草,影响了许多本地植物的生长。研究主要集中在1 年生草本植物[20-21],鲜有关注鬼针草对多年生小灌木的影响。考虑到灌木在植物群落中的重要性,研究这种植物相互作用,能补充鬼针草的入侵策略。因此,本研究选择入侵植物鬼针草和黔中地区常见本地植物醉鱼草(Buddleja lindleyana)为对象[22-23],探讨2 种植物在不同水分处理下,植物地上和地下形态、生物量特征对种间相互作用的响应,回答以下问题:(1)不同水分条件下,外来植物鬼针草与本地植物醉鱼草的种间关系。(2)干旱条件下,种间作用对鬼针草根、茎、叶特征的影响。为理解鬼针草与本地植物的种间关系和相关生长研究提供理论依据。
1 材料与方法
1.1 试验材料与设计
鬼针草为菊科(Asteraceae),鬼针草属(Bidens),1 年生入侵草本植物。醉鱼草为玄参科(Scrophulariaceae),醉鱼草属(Buddleja),多年生本地小灌木。2 种植物多分布于沟边、荒坡的灌木丛中,分布范围存在重叠,常与草本和灌木伴生,其中以禾本科及菊科植物居多[22-24]。
试验在贵州大学温室大棚中进行(26°27′13″N,106°40′19″E)。鬼针草和醉鱼草种子于2020 年6~8 月采集于贵州省贵阳市花溪区花溪水库附近,置于冰箱中5 ℃保存。2021年6月开始催芽,出苗后2 周内将醉鱼草和鬼针草幼苗各1 株移植于1个花盆中(直径20 cm,高17 cm),缓苗4周,于7 月26 日开始试验,11 月14 日结束试验。为减少土壤板结现象,保证根系取样完整,用均匀混合的无菌珍珠岩、石英砂和蛭石(体积比1∶1∶3)育苗[25]。测量花盆的饱和含水量,记录并称取20 盆配方相同的花盆m1,分别向所有花盆注水,直至盆底渗水且表面无积水,记录质量m2,饱和含水量为m0=m2-m1[26],即(1 542.71±35.27)g,n=20。缓 苗2周后对每盆植物统一施加基肥,养分条件为全氮1.030 g·kg-1,全磷0.099 g·kg-1,全钾3.860 g·kg-1,速效磷4.180 mg·kg-1,速效钾24.124 mg·kg-1。出苗前40 d 保持水分90%~100%的饱和含水量的条件,40 d 后开始水分处理,持续50 d。分别在生长40 d和90 d时取样并测量。
试验采用裂区设计,处理包括水分处理和种植处理。水分用称量法控制,包括对照和干旱;种植处理包括种间组和挡板组。水分处理参考宋会兴等[27]、王青宁等[28]对鬼针草和醉鱼草干旱胁迫的研究,设置水分条件为90%~100%的饱和含水量作为对照组,干旱条件为30%~35%的饱和含水量。每2天称量补水。种植处理中,种间组在同一花盆种植鬼针草和醉鱼草各1 株,株间距2 cm。挡板组用无色透明塑料挡板(长40 cm,宽20 cm)将花盆均匀分割为两部分,鬼针草和醉鱼草各1株分别种植于挡板两侧。挡板地上部分长23 cm,将花盆地上和地下部分分隔。每种处理每个物种10个重复。
1.2 数据收集与分析
生长40 d 和90 d 时取样,40 d 收获挡板组和种间组共20 盆,40 株,作为植物早期生长大小的估计,90 d 收获干旱和对照处理下的种间组、对照组共40 盆,80 株。测量每株植物总叶面积、整株分成根、茎、叶3 部分,于烘箱中60 ℃烘干至恒质量。计算比叶面积,总生物量和根冠比。每次取样将根系置于根系扫描仪中获取图像,用WinRhizo 根系分析软件获得植物根长、根平均直径、根表面积和根尖数量等形态参数。使用相对生长率(RGR,%)反映水分处理对植物生长的影响[29],计算公式如下:
式中:Y表示植物90 d 取样时各特征的观测值,X表示植物40 d 取样时各特征的观测值的平均值。
数据分析使用SPSS 20.0,绘图使用Origin 2018。各组试验处理的数据均符合正态分布和方差齐性。对数据进行物种、水分处理和种间作用的三因素方差分析(Three-ANOVA);使用单因素方差分析(One-ANOVA)不同水分条件或种间作用对鬼针草和醉鱼草各特征值相对生长的影响,多重比较用最小显著差异法(LSD)。
采用相对邻株效应指数(RNE)[30-31]计算90 d取样时鬼针草和醉鱼草种间作用的结果,计算公式如下:
式中:BO表示挡板组处理下10 株植物的茎、叶、根生物量或总生物量的平均值,BW表示种间处理下10株植物生物量平均值(同上)。
RNE 的范围从-1 到+1,负值表示促进,正值表示竞争,以此来量化鬼针草和醉鱼草之间的竞争与促进关系[32]。
2 结果与分析
2.1 种间相互作用的结果
40 d 取样时鬼针草和醉鱼草总生物量的RNE值分别为0.211、0.076,均表现为竞争,且鬼针草受到的竞争强于醉鱼草(表1)。鬼针草根、茎、叶的RNE 值均为正,种间相互作用表现为竞争;醉鱼草叶和根RNE 值为正值,表现出竞争,仅茎RNE 值为负,表现为促进。90 d 取样时干旱处理下鬼针草总生物量RNE 值为-0.291,种间关系表现为促进,而醉鱼草RNE值为0.067,表现为竞争。鬼针草的种间关系由竞争变为促进,而醉鱼草种间关系在对照处理下RNE值为负,表现出促进,而干旱处理RNE 值却为正,表现出竞争。鬼针草的根、茎、叶RNE 值均为负,表现为促进;2 种水分条件下醉鱼草茎RNE值为负,表现为促进,对照处理下叶RNE值为正,表现为竞争,而干旱下其值为负,表现为促进。醉鱼草根的种间关系与叶的结果相反。
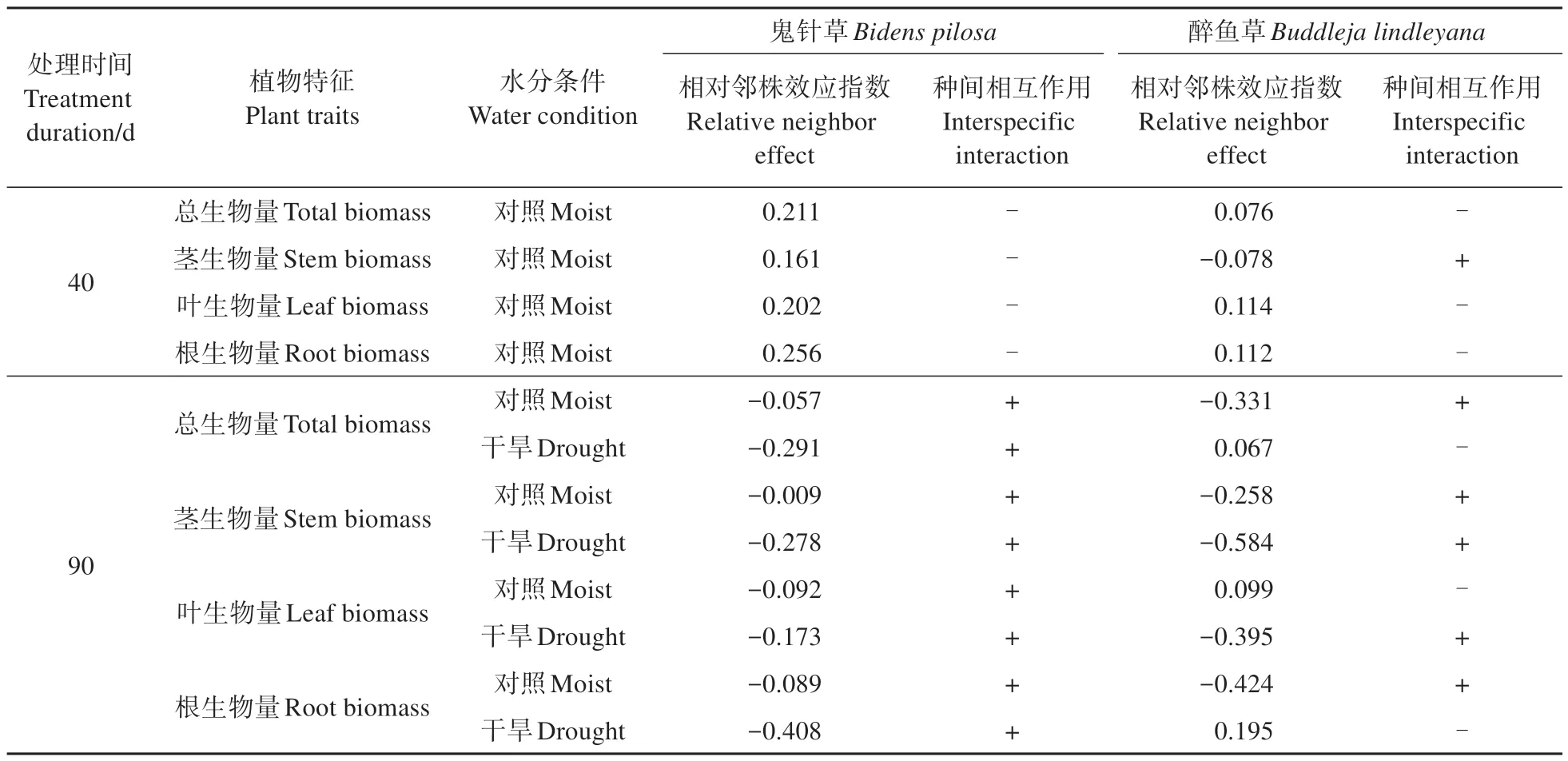
表1 不同水分条件下鬼针草和醉鱼草的种间相互作用Table 1 Interspecific interaction between Bidens pilosa and Buddleja lindleyana under different water conditions
2.2 水分处理对植物各特征相对生长的影响
水分处理对2 个物种叶面积和比叶面积都有显著影响(P<0.05,表2)。与对照相比,干旱处理显著降低了鬼针草的叶面积(P<0.05,图1A),并降低挡板组下醉鱼草的叶面积和比叶面积(P<0.05)。与对照组相比,干旱降低鬼针草种间组下的比叶面积(P<0.05,图1B)。
水分处理对2个物种的总生物量、叶生物量和根生物量的相对生长率都有显著影响(P<0.05,表1)。与对照组相比,干旱处理显著降低鬼针草的总生物量和叶生物量(P<0.05,图2A,E);与对照相比,干旱处理显著降低醉鱼草在种间组的总生物量、根生物量和根冠比(P<0.05,图2A,B,E),干旱处理显著降低醉鱼草在挡板组的茎和叶生物量(P<0.05,图2C,D),却显著提高其根冠比(P<0.05,图2B)。
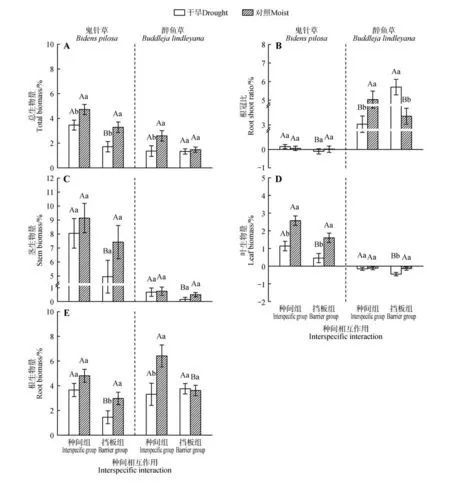
图2 水分处理和种间作用对总生物量(A)、根冠比(B)、茎(C)、叶(D)、根(E)生物量相对生长率的影响Fig.2 Effects of water treatment and interspecific interaction on the relative growth rate of total biomass(A),rootshoot ratio(B),stem(C),leaf(D),root(E)biomass.
2.3 相同水分条件下种间相互作用对植物的影响
种间相互作用对2 个物种的叶面积和比叶面积有显著影响(P<0.01,表2)。与挡板组相比,种间相互作用显著降低干旱下鬼针草的叶面积和比叶面积(P<0.05,图1A,B);与挡板组相比,种间相互作用显著降低对照处理下醉鱼草的叶面积和比叶面积(P<0.05)。
种间作用对2 个物种的各生物量的相对生长率均有显著影响(P<0.05,表1)。与挡板组相比,种间相互作用显著提高干旱下鬼针草的总生物量、茎、根、叶生物量和根冠比(P<0.05,图2);与挡板组相比,种间相互作用显著提高干旱条件下醉鱼草茎和叶生物量(P<0.05),却显著降低其根冠比(P<0.05)。与挡板组相比,种间相互作用显著提高对照组水分下醉鱼草的根冠比和根生物量(P<0.05)。与挡板组相比,种间相互作用显著提高鬼针草和醉鱼草在干旱和对照组水分条件下的根长、根表面积和根尖数量(表3)。
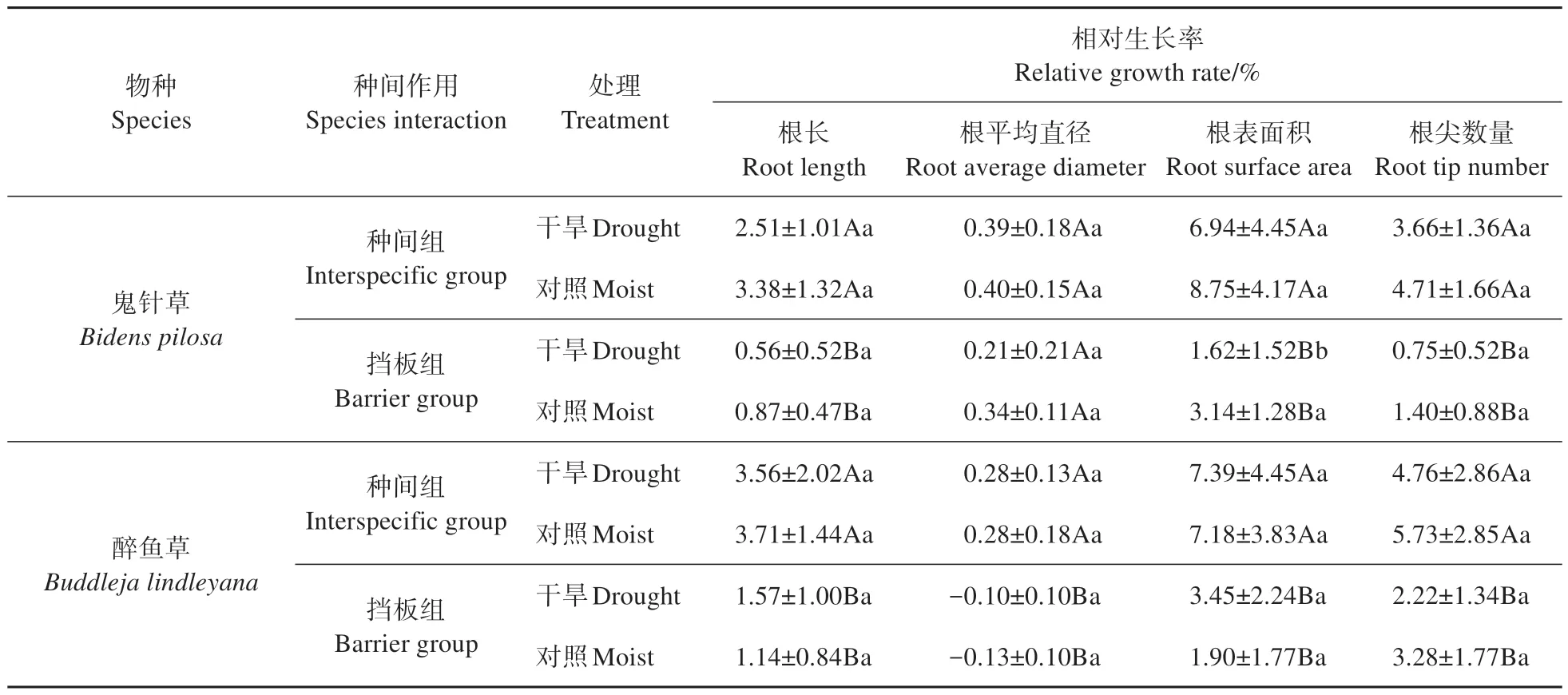
表3 水分处理和种间作用对根系生长的影响Table 3 Effects of water treatment and interspecific interaction on root growth
3 讨论
3.1 干旱条件对种间关系的影响
极端环境下植物的种间关系更倾向于促进作用,可以缓解不利环境对植物的影响[33]。本研究发现,第一阶段鬼针草种间关系为竞争,第二阶段干旱处理后鬼针草的种间关系表现出明显的促进,根受到的促进作用最大;而醉鱼草第一阶段种间关系为弱竞争,第二阶段干旱处理下仍表现为竞争,但茎和叶生物量受到促进作用。对于鬼针草,干旱可能改变其种间关系,并且能增加该物种受到的促进作用,虽然对照组RNE指数为负,但其数值较小表现为弱促进,可能是生长阶段不同导致鬼针草种间关系的变化,因为幼苗生长的早期阶段植株较小,与邻株植物发生的相互作用较少,即种间关系较弱,随着植株生长,根系间的觅食和叶片争夺光照会增强种间的相互作用。对于醉鱼草,干旱虽然对叶和茎有促进作用,但会增加根系的竞争,说明干旱会影响醉鱼草根、茎、叶的竞争或促进,而鬼针草的种间关系较为稳定。一方面说明,醉鱼草竞争与促进的平衡可能与水分条件有关,并且干旱处理增加了根系竞争的同时,可能掩盖了茎和叶受到的促进效果。另一方面说明,不同物种地上和地下的种间关系可能受水分的影响。2 个物种根生物量的种间关系与总生物量相同,这种现象与meta-analysis 5 项地上与地下竞争的研究结果[7]相同,表明植物相互作用主要是发生于根部,地上部分的作用较弱[34]。所以试验中的根系对水分变化响应敏感,容易受到干旱的影响从而改变其种间关系。
3.2 干旱条件下种间相互作用对植物生长的影响
种间相互作用可能限制了植物对非生物环境胁迫的反应,如植物会通过表型的调整尽可能地提高资源的利用效率,而种间相互作用可能改变这种环境下植物形态和生物量特征[35]。本研究表明,与挡板组相比,种间相互作用增加了干旱条件下鬼针草根、茎、叶生物量的生长和根冠比(图2),以及根长、根表面积和根尖数量(表3);虽然鬼针草各生物量表现为促进(表1),但叶片形态上的表现存在竞争,如与挡板组相比,种间作用降低其叶面积和比叶面积(图1),表明叶生物量受到的促进作用并不意味着叶面积会增加。此外植物面对低环境资源时往往采取保守的生长策略,故会减少新叶的生长,从而减少叶片特征[36]。这种生物量与形态特征表现出相反的种间关系,这可能是植物同时适应干旱和种间相互作用的策略。对于醉鱼草,种间作用增加了干旱条件下茎、叶生物量和根长、根平均直径、根表面积和根尖数量,并降低了根冠比,一般情况下,植物会增加根冠比来响应干旱[37],而醉鱼草根系可能受到了强烈的竞争抑制其地下生物量的分配。
3.3 入侵植物和本地植物
有研究表明鬼针草对1 年生杂草群落有显著影响[38],但对于多年生灌木的研究较少。本试验中,种间相互作用能促进鬼针草生物量的积累,并降低本地种醉鱼草的比叶面积和根冠比,说明野外环境下鬼针草可能对醉鱼草生长产生影响。此外,入侵植物具有较强的适应能力或表型可塑性[39-40]。鬼针草入侵策略主要是增加地上部分的生长,这种表现与入侵植物加拿大一枝黄花(Solidago canadensis)相同,具有快速的生长能力,迅速占据本地植物的生境[41-42]。另外,鬼针草增加枝条向外扩展的能力[43],便于枝条末端种子的扩散,利于后代扩散的亲本投资[44]。而醉鱼草增加茎生物量生长,通过调整根冠比来适应水分变化。比较2种水分条件,入侵植物鬼针草在干旱环境受到的促进效果最大,表明干旱或残酷生境下,与醉鱼草建立的种间关系,能提高鬼针草的表现和入侵性。
4 结论
干旱能影响2个物种的种间关系,鬼针草和醉鱼草受到的影响不同。这种种间作用可能会促进干旱地区鬼针草的进一步的入侵和扩散,使鬼针草在干旱环境具有优势。鬼针草主要影响醉鱼草的叶片生长和根冠比。应当持续关注不同环境因子下,鬼针草与其他植物建立的种间相互作用,从而为了解该物种入侵提供理论依据。
猜你喜欢
中学生数理化·自主招生(2023年3期)2023-03-17 02:35:02
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
电站辅机(2021年4期)2021-03-29 01:16:52
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
蔬菜(2018年5期)2018-05-17 06:56:36
现代农业科技(2017年16期)2017-09-22 14:47:46
杂草学报(2015年2期)2016-01-04 14:57:55
江苏农业科学(2014年1期)2014-07-18 21:21:34
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:02
