
4种光响应曲线模型对3种高寒草甸植物的实用性分析
2018-07-28 08:18,,,,,
草地学报 2018年2期
, , , , ,
(1. 西北师范大学地理与环境科学学院,甘肃 兰州 730070; 2. 兰州城市学院城市生态与环境生物技术中心, 甘肃 兰州 730070; 3. 兰州大学生命科学学院,甘肃 兰州 730070)
光合作用是绿色植物利用光能将水分和二氧化碳转化为有机物质并释放氧气的过程[1],植物对于光环境变化的适应能力决定了它的物种丰度和分布[2]。金露梅(PotentillafruticosaL.)、野草莓(Fragariavesca)和酸模(RumexacetosaL.)是广泛分布在甘肃西南部积石山山麓高山草甸坡地上的优势草本植物;积石山黄草坪草场是积石山县主要牧场之一,近年来由于过度载畜、旅游野炊等人为活动的影响,使该地区植被破坏,草场退化,生态环境异常脆弱。开展植物光响应机制的研究,能揭示草地植物的生理生态学机制和生长状况。对于合理适量的利用和保护草地,恢复受损草地生态系统具有非常重要的现实和理论意义。
国内外学者对高寒草地植物的光合作用做了很多研究,如郭亚奇[3]等探讨了土壤水分、温度和CO2浓度升高对藏北高寒草地紫花针茅(Stipapurpurea)光合作用的影响;孙英[4]等对祁连山东段高寒地区草地优势植物旺盛生长期的光合特征进行了研究;闫霜[5]揭示了青藏高原东缘不同氮处理水平下异针茅(Stipaaliena)叶片光合作用的响应规律;肖红[6]等通过模拟牦牛、藏羊践踏,研究天祝高寒草地阴山扁蓿豆(Medicagorutheniavar.inschanicus)单叶面积、叶绿素含量、叶片气体交换参数变化,揭示了降水的响应规律;Cui Xiaoyong[7]等探索了青藏高原高寒草甸和湿地两种苔藓植物的动态光合作用;LI Meiling[8]等专家解释了温度增强对沙柳河高寒草甸群落光合作用日变化的影响。已有研究主要侧重于生态因子对高寒草地植物光合特性及其日变化的影响。通过光响应曲线的拟合揭示出环境因子对光合作用的实用性分析在该地区未见报道。
光响应曲线是探究植物光合特性的主要方法之一[9],通过光响应曲线可计算出植物的光饱和点、光补偿点、最大净光合效率、初始量子效率及暗呼吸速率等光合参数。因此,研究植物的光响应曲线有助于解释环境因子与植物光合产物积累的关系。常用的模型有直角双曲线模型、直角双曲线修正模型、指数模型和非直角双曲线模型等。同一种植物用不同模型拟合出的光响应特征参数存在差异,为保证光合特征参数的准确性,了解植物的光合特性,需对不同的光响应模型进行比较。本研究拟选用4种模型拟合3种植物的光合-光响应曲线,并通过比较相关系数、均方根误差、平均绝对误差及拟合值和实测值的相对误差之和得出光响应参数的最适模型,揭示金露梅(PotentillafruticosaL.)、野草莓(Fragariavesca)和酸模(RumexacetosaL.)在强光低温环境的光合生理机制和生存适应性,为高寒草甸的利用和退化恢复提供参考。
1 材料与方法
1.1 供试材料
2017年6月在甘肃西南部积石山保安族东乡族撒拉族自治县积石山山麓高山草甸黄草坪地坡地(海拔2 734 m,35°42' N,102°47' E),属典型的大陆性季风气候区,年均温度7.2℃,最高气温12.5℃,最低气温3.1℃,年均降水606 mm,年均日照时数2 323 h,无霜期130 d,太阳总辐射量为131.34 KCal·cm-2,土壤为高山草甸土,pH为6.8~7,土壤有机质含量高。选择该地区金露梅、野草莓和酸模的成熟植株外围向阳叶片为待测对象。
1.2 测量方法
1.2.1光响应曲线的测定 试验在6月中旬晴朗天气的上午9:00-12:00进行。采用LI-6400便携式光合仪对植物叶片的净光合速率(Pn)进行测定,每种植物随机选取3株长势较一致的成熟植株,每株挑选外围向阳的叶片进行测量,3次重复。测量前将待测叶片在800 μmol·m-2·s-1光强下进行光合诱导。使用开放气路,设定流速为500 μmol·s-1,CO2浓度为400 μmol·mol-1。用仪器配备的红、蓝人工光源(LI6400-02BLED),设定光合有效辐射强度(PAR)由高到低为2 000 μmol·m-2·s-1、1 600 μmol·m-2·s-1、1 200 μmol·m-2·s-1、800 μmol·m-2·s-1、400 μmol·m-2·s-1、0 μmol·m-2·s-1。测定时最大等待时间设为2 min,每次测定后都要进行匹配,测量过程中也要对仪器进行自动匹配操作。叶面积指数的测量采用仪器叶面积指数仪LAI-2000进行测量。
根据实测净光合速率随光强的变化情况,即实测光响应曲线,通过目视判断获得最大净光合速率(Pnmax、μmol·m-2·s-1)、光补偿点(LCP、μmol·m-2·s-1,Pn=0时的光合有效辐射)、光饱和点(LSP、μmol·m-2·s-1)和暗呼吸速率(Rd、μmol·m-2·s-1,光合有效辐射为0时的Pn)的观测值。
1.2.2光响应曲线拟合模型 光响应曲线的拟合模型有许多,广泛应用于植物生理生态等方面研究的有指数模型、正切函数曲线模型、二项式回归模型、非直角双曲线模型、直角双曲线模型、Michealis-Menten模型、Farquhar生化模型和直角双曲线修正模型[10]。直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型是其最常用的几个模型,虽有研究表明个别植物拟合效果最好的并非直角双曲线修正模型[11-13],但大多研究表明直角双曲线修正模型在不同生境下的拟合效果最好[1,14-16],而对该地区植物拟合最适用的模型是否是直角双曲线修正模型尚不明确。本文选择4种最常用的直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型进行光响应曲线的拟合[17-20]:
(1)
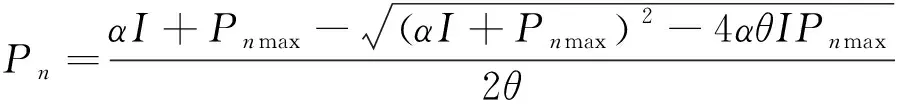
(2)
(3)
(4)
式中:Pn为净光合速率,α为初始量子效率,I为光合有效辐射,Pnmax为最大净光合速率,Rd为暗呼吸速率。θ表示光响应曲线弯曲程度的参数,取值范围为0<θ<1。β、γ是修正系数;β为光抑制项;γ为光饱和项,其值为α/Pnmax。
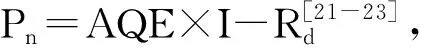
在模型(3)中,光饱和点是0.99Pnmax对应的光照强度,光补偿点为Pn=0时的光强[24]。
在模型(4)中,Pnmax、LSP、LCP的计算公式为:
(4a)
(4b)
(4c)
为了更好地检验4种模型的拟合度,除相关系数(R2)外,另选2种指标对模型的拟合效果进行评价:均方根误差(RMSE)和平均绝对误差(MAE)[12,15]:
(5)
(6)

1.3 数据分析
用Excel软件对数据进行初步处理,Origin 8.0作图,SPSS 21.0统计软件对3种植物的光响应曲线进行直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型的拟合分析。
2 结果与分析
2.1 光合-光响应曲线的拟合及比较
直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型拟合三种植物的光响应曲线,均得到了较好的拟合效果。图1表明,各拟合曲线的坡度都是先陡后缓,光强小于400 μmol·m-2·s-1时,随光照强度的增加净光合速率增加快速;当金露梅和酸模的光强大于400 μmol·m-2·s-1小于1 200 μmol·m-2·s-1,而野草莓光强大于400 μmol·m-2·s-1小于800 μmol·m-2·s-1时,随光照强度的增加净光合速率增加变缓;金露梅和酸模的光强大于1 200 μmol·m-2·s-1,野草莓光强大于800 μmol·m-2·s-1时,坡度变平缓,净光合速率基本不变,没有发生下降的现象。对3种植物的光响应曲线选用4种模型进行拟合,通过拟合方程得到的相关系数R2可知,4种模型对金露梅光响应曲线的拟合程度大小是:直角双曲线修正模型(0.9995)、非直角双曲线模型(0.9990)、指数模型(0.9988)、直角双曲线模型(0.9983);野草莓的拟合模型中直角双曲线修正模型、直角双曲线模型和非直角双曲线模型的R2值(0.99935)相同,都大于指数模型(0.99800);对酸模光响应曲线的拟合程度大小是:非直角双曲线模型(0.99997)、直角双曲线修正模型(0.99994)、直角双曲线模型(0.99958)、指数模型(0.99904)(表1)。R2越大越接近1,说明拟合的程度越高,但并不能说明拟合效果就最好。因此,还要对光响应曲线的拟合参数进行分析。
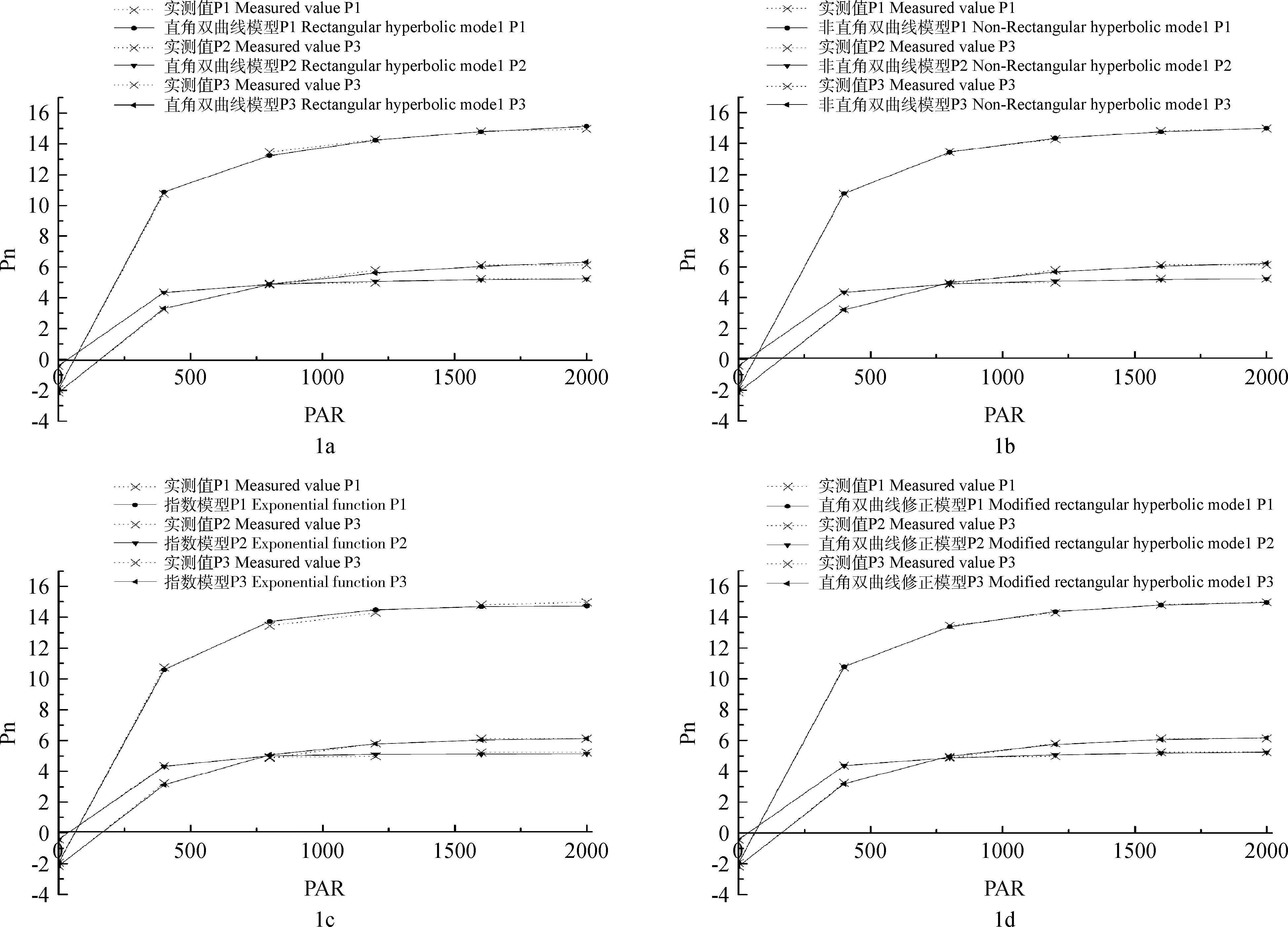
图1 四种模型对3种植物的光合光响应曲线的模拟Fig.1 Four models were used to simulate the photosynthetic response curve of three plants注:1a:直角双曲线模型;1b:非直角双曲线模型;1c:指数模型;1d:直角双曲线修正模型 Note:1a: Rectangular hyperbolic model;1b: Non-rectangular hyperbolic model;1c: Exponential function;1d: Modified rectangular hyperbolic model
2.2 不同模型拟合的光响应参数的比较分析
由表1可知,4种模型拟合的酸模最大净光合速率顺序为:直角双曲线修正模型16.263 μmol·m-2·s-1、指数模型16.667 μmol·m-2·s-1、非直角双曲线模型17.847 μmol·m-2·s-1、直角双曲线模型18.603 μmol·m-2·s-1,4种模型对酸模Pnmax的拟合值均大于实测值14.965 μmol·m-2·s-1,尤其是直角双曲线模型对Pnmax的拟合值18.603 μmol·m-2·s-1,而直角双曲线修正模型的拟合值最接近于实测值;4种模型对金露梅和野草莓的最大净光合速率的拟合情况与酸模相同,都大于实测值,且直角和非直角双曲线模型对金露梅和野草莓的拟合值9.838 μmol·m-2·s-1、9.21 μmol·m-2·s-1, 5.923 μmol·m-2·s-1、5.901 μmol·m-2·s-1都远大于实测值6.12 μmol·m-2·s-1, 5.228 μmol·m-2·s-1,直角双曲线修正模型的拟合值6.33 μmol·m-2·s-1、5.302 μmol·m-2·s-1最接近于实测值6.12 μmol·m-2·s-1、5.228 μmol·m-2·s-1。4种模型对暗呼吸速率Rd的拟合中,酸模植物直角双曲线修正模型和非直角双曲线模型的拟合值1.931 μmol·m-2·s-1、1.930 μmol·m-2·s-1都最接近实测值1.930 μmol·m-2·s-1,直角双曲线模型和指数模型的拟合值1.935 μmol·m-2·s-1、1.909 μmol·m-2·s-1均与实测值相差较大;野草莓暗呼吸速率的指数模型拟合值0.422 μmol·m-2·s-1略小于实测值0.425 μmol·m-2·s-1,其他三种模型拟合值与实测值相同;金露梅暗呼吸速率直角双曲线修正模型和非直角双曲线模型的拟合值2.149 μmol·m-2·s-1、2.152 μmol·m-2·s-1接近于实测值2.15 μmol·m-2·s-1,直角双曲线模型和指数模型的拟合值2.161 μmol·m-2·s-1、2.133 μmol·m-2·s-1与实测值相差较大。对比4种模型的光补偿点LCP和光饱和点LSP的拟合值可知,4种模型对酸模植物的LCP拟合值为:直角双曲线模型18.786 μmol·m-2·s-1、非直角双曲线模型35.09 μmol·m-2·s-1、指数模型34.956 μmol·m-2·s-1、直角双曲线修正模型21.943 μmol·m-2·s-1,直角双曲线修正模型最接近于实测值27.90 μmol·m-2·s-1;直角双曲线模型、非直角双曲线模型和指数模型的LSP拟合值199.398 μmol·m-2·s-1、359.582 μmol·m-2·s-1、648.923 μmol·m-2·s-1都远小于实测值,直角双曲线修正模型对LSP的拟合值2 389.806 μmol·m-2·s-1最接近于实测值2 000 μmol·m-2·s-1;对野草莓的LCP拟合值为:直角双曲线模型6.746 μmol·m-2·s-1、非直角双曲线模型8.673 μmol·m-2·s-1、指数模型16.256 μmol·m-2·s-1和直角双曲线修正模型6.746 μmol·m-2·s-1,直角双曲线模型和直角双曲线修正模型最接近于实测值6.90 μmol·m-2·s-1;直角双曲线修正模型的LSP拟合值1 661.920 μmol·m-2·s-1最接近于实测值1 600 μmol·m-2·s-1,其他三种模型都远小于实测值;拟合的金露梅叶片的LCP值为:直角双曲线模型69.71 μmol·m-2·s-1、非直角双曲线模型102.48 μmol·m-2·s-1、指数模型117.35 μmol·m-2·s-1和直角双曲线修正模型85.96 μmol·m-2·s-1,直角双曲线修正模型最接近于实测值96.90 μmol·m-2·s-1,直角双曲线修正模型拟合的LSP是2 111.24 μmol·m-2·s-1也最接近于实测值2 000 μmol·m-2·s-1,其他三种模型都远小于实测值。综上,对酸模、野草莓和金露梅叶片的拟合模型中直角双曲线修正模型的拟合结果均优于其它三种模型。

表1 不同光合光响应模型拟合的光合参数及其与实测值的比较Table 1 Comparison of photosynthetic parameters of different photosynthetic light response models and their comparison with measured values
注:P1为酸模,P2为野草莓,P3为金露梅
Note: P1 for theRumexacetosaL., P2 for theFragariavesca, and P3 for thePotentillafruticosaL.
2.3 不同模型拟合优度比较
根据表2均方根误差和平均绝对误差值可知,酸模直角双曲线模型和指数模型的RMSE(0.1228、0.1860)和MAE(0.0973、0.1673)值均较大;非直角双曲线模型和直角双曲线修正模型的RMSE(0.0324、0.0472)和MAE(0.0240、0.0338)值相差较小,表明两者拟合度接近,都优于直角双曲线模型和指数模型。野草莓除指数模型的RMSE和MAE的值较大外,其它三种模型的RMSE(0.0520、0.0505、0.0520)和MAE(0.0367、0.035、0.0367)的值相差很小,表明这三种模型的拟合度接近。金露梅直角双曲线修正模型的RMSE(0.067)和MAE(0.057)的值最低,其它三种模型的RMSE(0.121、0.102、0.095)和MAE(0.093、0.077、0.082)值都远高于直角双曲线修正模型,经过T检验得出3种植物的RMSE与MAE均在95%的置信区间内有显著性(P<0.05)。分析不同光合有效辐射下净光合速率的实测值和4种模型的估计值,表现出4种模型拟合的光合光响应曲线与实测值有着基本一致的变化趋势(表3)。对于4种模型拟合值的相对误差之和,酸模植物指数模型和直角双曲线模型的相对误差之和都偏大,直角双曲线修正模型和非直角双曲线模型的相对误差之和均偏小且较接近,表明直角修正模型和非直角双曲线模型拟合结果更偏向于实测值;野草莓除指数模型的相对误差之和(0.094)偏大外,另三种模型的相对误差之和都偏小且非常相近(0.044、0.042、0.044);金露梅的相对误差之和最小的是直角双曲线修正模型(0.068),其它模型的值都远大于直角双曲线修正模型。结合表1,3种植物的拟合效果最好的都是直角双曲线修正模型。
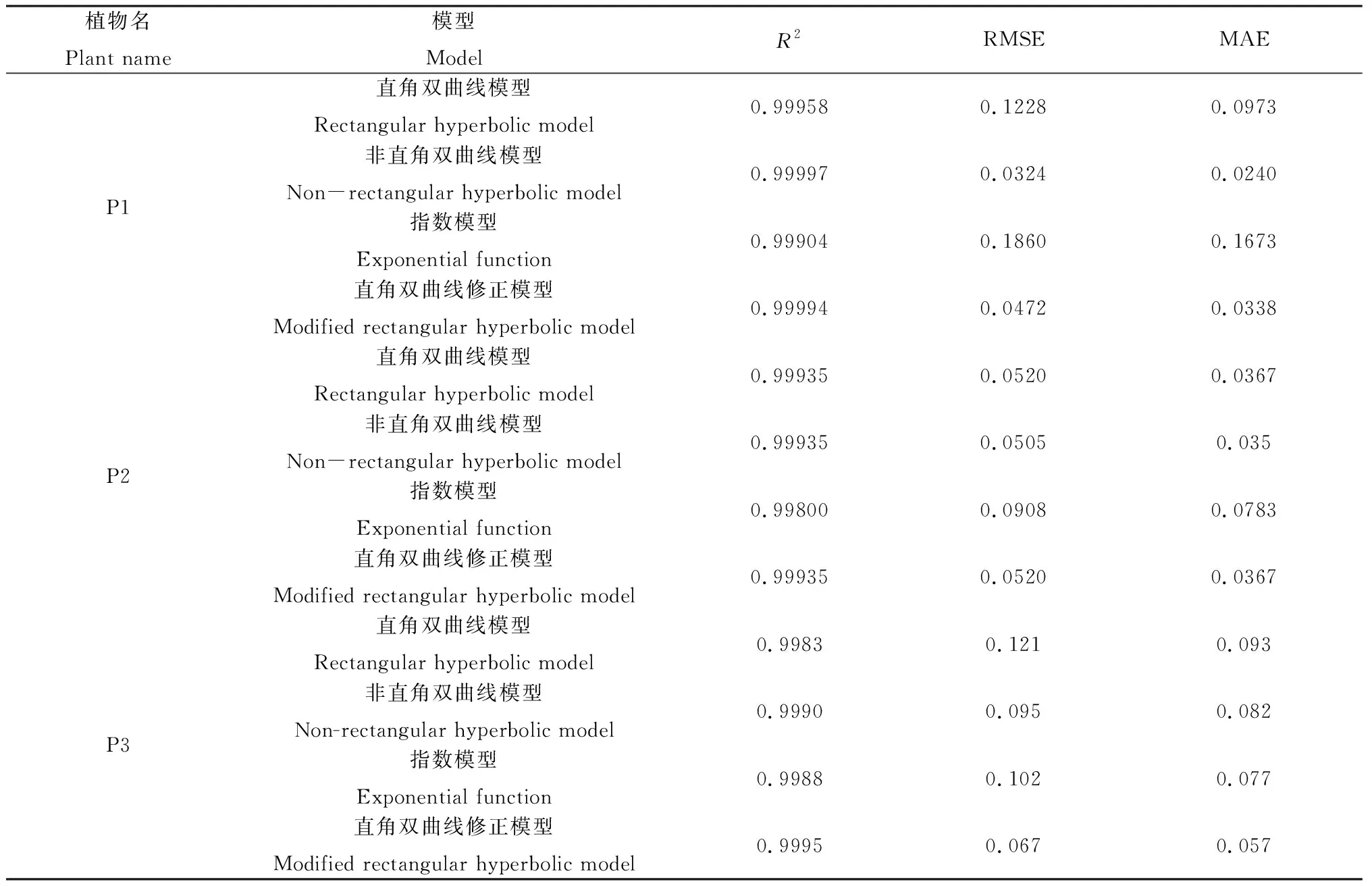
表2 由相关系数(R2)、均方根误差(RMSE)和平均绝对误差(MAE)表示的4种模型的拟合优度Table 2 The fit of the four models represented by correlation coefficient (R2), root mean square error (RMSE) and mean absolute error (MAE)
注:P1为酸模,P2为野草莓,P3为金露梅
Note: P1 for theRumexacetosaL, P2 for theFragariavesca, and P3 for thePotentillafruticosaL.

表3 不同光合有效辐射下净光合速率的实测值和拟合值相对误差之和的比较Table 3 The comparison between the measured value of net photosynthetic rate and the relative error of the fitting value in different photosynthetic effective radiation
3 讨论
光合-光响应曲线模型是研究植物生理的重要工具,为植物光合特性对环境因子的响应提供科学依据。不同植物有其最适的光合光响应曲线模型。直角双曲线模型和非直角双曲线模型是一条渐近线,无极值,在实际应用过程中,会出现饱和光强远小于实测值;若光强达到饱和后净光合速率不降低,则估测的Pnmax值必大于实测值[25],这与本实验结果一致。指数模型对于本文中3种植物叶片的拟合得到的Pnmax较为接近实测值,但人为干扰较大,生物学意义不明确。直角双曲线修正模型对于3种植物叶片Pnmax的拟合值均最接近实测值,而许多研究也表明了直角双曲线修正模型对大多数植物光响应曲线的拟合有较好的适应性[9,13,26-27]。在直角双曲线修正模型中,β 为光抑制项,γ 值一定时,β 值越大,光响应曲线弯曲程度就变大,植物受光抑制就越强,图1中3种植物叶片光响应曲线的弯曲程度不同,野草莓叶片光响应曲线的弯曲程度较大,植物易受光抑制,其饱和光强相对较低(1600 μmol·m-2·s-1),金露梅和酸模叶片光响应曲线弯曲程度较小,植物不易发生光抑制,饱和光强则相对较高(2000 μmol·m-2·s-1)。当β 一定时,γ 值越小,表示光响应曲线弯曲程度就越小,不易发生光饱和现象,对应饱和光强的值就大。3种植物叶片光响应曲线的弯曲程度证明了其γ 和β 的生物学意义,也体现了该模型相对于其他模型更广泛的适应性。直角双曲线修正模型方程是存在极值的,其拟合曲线与实测曲线符合度很高,本研究与这一结果相符。且通过MRSE和MAE评价指标结合R2以及比较4种模型在不同光照强度下净光合速率的估计值和实测值的相对误差之和检验其结果,也表明直角双曲线修正模型的拟合度高。
有研究表明通过直角双曲线修正模型拟合研究比较电子传递速率和光合作用对光响应曲线的饱和光强可判断植物的电子传递速率与碳同化达到最大值的时间是否一致[28],可为进一步研究植物的光合特性和植物对逆境的响应提供一种有效的模拟工具。直角双曲线修正模型不仅能直接得出所求参数,且得到的Pnmax和LSP最接近于实测值,对3种植物叶片的高光强和低光强部分均有较好的拟合效果,其模型参数也有较明确的生物学意义,体现了其广泛的适应性。但是否对高寒阴湿环境下的所有植物都能够适应,还需进一步研究。
3种植物叶片的光合能力存在一定差异,是其生理、形态和生化等的遗传特性的反应及对高寒阴湿地适应的结果。在一定条件下,植物的最大光合能力决定于电子传递的速率。3种植物叶片最大净光合速率的大小为:酸模>金露梅>野草莓,说明酸模叶片维持了较高的PSII电子传递速率。LSP和LCP是植物利用强弱光能力的大小[29-30],三种植物的LSP由大到小为:酸模>金露梅>野草莓;LCP由大到小顺序为:金露梅>酸模>野草莓,说明酸模忍耐强光的能力最强,能适应更多的强光环境,不易发生光抑制;野草莓的LCP值最低,说明它适应弱光的能力较强。暗呼吸速率最小的是野草莓(0.425),说明在无光的条件下,野草莓消耗的光合产物最少。表观量子效率反应叶片利用光能的情况,尤其是对弱光的利用,值越大说明植物利用弱光的能力越强,植物转化吸收光能的色素蛋白越多[31]。3种植物中表观量子效率最大的是酸模,因此其对光响应最敏感,对弱光的利用率也最高。从叶片形态来讲,酸模叶片最大,叶面积指数为5.18,金露梅为簇状叶片,叶面积指数为3.32,野草莓叶面积指数为2.28,叶面积大的植物,叶肉细胞叶绿体数目也更多,能够截获更多的光资源用于光合作用,从而积累更多的光合产物用于自身的生长繁殖。
4 结论
本文采用的4种模型对黄草坪3种植物叶片光合-光响应曲线的拟合,其最优模型是直角双曲线修正模型,即直角双曲线修正模型对卵状披针形叶片、倒披针形叶片和长圆状披针形叶片的拟合均最佳。酸模对环境的适应能力最强,金露梅次之,野草莓最弱。该地区属于高寒阴湿地区,野草莓消耗光合产物最少,金露梅和酸模均具有较高的光能转换效率。研究结论为积石山地区生态保护提供一定的理论参考。
猜你喜欢
环境保护与循环经济(2021年7期)2021-11-02
哈尔滨轴承(2020年1期)2020-11-03
中国奶牛(2019年10期)2019-10-28
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
电子制作(2018年23期)2018-12-26
中学生数理化·七年级数学人教版(2017年9期)2017-08-15
中学生数理化(高中版.高二数学)(2017年1期)2017-04-16
中学数学杂志(2015年9期)2015-01-01
