
姜黄素调控Wnt信号通路治疗脊髓损伤研究进展
2018-07-18 09:23:34张宋欧徐璐瑶周静莹袁佳莹刘学红
中国运动医学杂志 2018年1期
张宋欧 徐璐瑶 周静莹 袁佳莹 刘学红
绍兴文理学院医学院(浙江绍兴 31200)
脊髓损伤(spinal cord injury,SCI)包括原发性损伤和继发性损伤,表现为不同程度的脊髓神经组织受损,轻者影响患者行为和功能,重者导致瘫痪,甚至死亡[1]。SCI发病率较高,美国约54/百万[2],中国约 23.7/百万[3]。原发性损伤难以逆转,故临床干预主要针对继发性损伤。由于原发性损伤的外力作用,而导致组织细胞受到不同程度的损伤,炎性信号通路被启动,损伤组织局部出现大量炎性细胞的集聚,在缺血缺氧的情况下,受损组织在炎性因子的作用下进一步受到损伤。继发性损伤,包括炎症反应、细胞凋亡、氧化应激、局部血管功能紊乱、轴突脱髓鞘、骨质丢失和胶质瘢痕形成等[4-5]。据报道Wnt、NF-κB、mTOR等信号通路在继发性脊髓损伤的病理过程均起到重要作用[6-7]。尤其是Wnt信号通路在神经系统发育和损伤修复过程中均发挥着重要的作用[7]。
1 Wnt信号通路
Wnt信号通路可分为两类:经典Wnt信号通路(Wnt/β-catenin信号通路),非经典信号通路(Wnt/PCP信号通路、Wnt/Ca+信号通路)。其中目前研究最多的是经典Wnt信号通路。Wnt/β-catenin信号通路由七部分组成:Wnt配体;跨膜受体(Frizzled,FZD);低密度脂蛋白受体相关的蛋白质家族(low-density lipid receptor-related protein,LRP);蓬乱蛋白(Disheveled,DVL);由Axin、糖原合成激酶(glycogen synthase kinase-3β,GSK-3β)、酪蛋白激酶1(casein kinase 1,CK1)和腺瘤性结肠息肉蛋白(adenomatous polyposis coli,APC)组成的β-catenin降解复合物;β-catenin;转录因子,T细胞促进因子和淋巴增强因子(T cell factor and lymphoid enhancer factor,TCF/LEF)[8]。
无Wnt信号刺激时,胞质内的Axin、GSK-3β、CK1和APC组成降解复合物结合β-catenin并使其磷酸化,随后β-catenin被泛素化和被蛋白酶体降解,这一过程维持β-catenin的低浓度状态。Wnt配体是Wnt信号通路激活过程的起点,与FZD、LRP5/6等跨膜受体结合,引起LRP-6的磷酸化和DVL结合到FZD、LRP蛋白上共同形成跨膜受体复合物,这一复合物的形成诱导了GSK-3β的磷酸化,造成降解复合物的解离,βcatenin在胞质内累积,并进入细胞核内,与TCF/LEF等转录因子产生相互作用,激活Wnt目标基因的表达,对细胞增殖、分化、炎症,神经发生和再生等产生一系列作用[9-10]。Wnt信号通路被认为与轴突发芽、神经发生和胚胎发育过程中的少突胶质前体细胞(Oligodendrocyte Precursor Cells,OPCs)迁移有关[11]。阿尔茨海默病、精神分裂症和帕金森病等神经退行性疾病都与Wnt信号通路调节有关。因此Wnt信号通路可以作为中枢神经系统疾病的潜在治疗靶点[12]。见图1。
2 Wnt信号通路与脊髓损伤
SCI后,Wnt信号通路的调控作用在继发性脊髓损伤的多个方面体现,如Wnt/β-catenin信号通路可抑制神经细胞凋亡与促进其再分化,抑制骨质丢失,Wnt/Ryk信号通路可促进损伤区域胶质瘢痕形成并抑制轴突再生[13-15]。
2.1 Wnt信号通路参与脊髓损伤后的炎症反应过程
炎症是细胞凋亡、轴突再生、胶质瘢痕形成等病理过程的基础,在SCI的病理过程中起到关键作用,激活Wnt信号通路可以抑制炎症。核转录因子(nuclear factor-κB,NF-κB)是一种重要的调控炎症的核因子,Wnt信号通路与NF-κB调控的信号通路存在交叉,βcatenin可以抑制NF-κB调控的一系列转录激活活动[16]。Guo等[17]认为β-catenin激活剂SB216763可以通过激活过氧化物酶体增殖物激活受体δ(Peroxisome proliferator-activated receptorδ,PPARδ)和丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)信号通路抑制炎症因子TNF-α和IL-1β。刺激Wnt3a可明显上调IL-10,下调TNF-α、IL-6和IL-1β[18]。糖皮质激素可以激活SCI后的Wnt/β-catenin信号通路,进而激活靶基因PPAR-γ,下调炎症因子IL-1β和TNF-α[19]。参见图1。
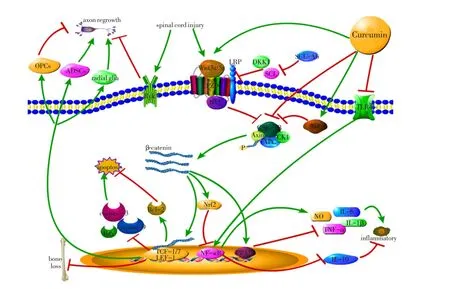
图1 姜黄素调控Wnt信号通路治疗脊髓损伤的作用机制
2.2 Wnt信号通路参与SCI后细胞凋亡过程
SCI后神经元凋亡会阻碍脊髓损伤后的功能恢复,抑制神经细胞凋亡和促进神经细胞再生对脊髓功能的恢复至关重要[20]。Wnt信号通路有利于SCI后神经细胞存活,激活Wnt通路可抑制SCI后神经细胞凋亡。Gao等[13]报道,辛伐他汀激活Wnt/β-catenin信号通路,促进LRP-6磷酸化,促进β-catenin表达,增加LEF-1和TCF-1mRNA表达,抑制β-catenin在脊髓神经元中的磷酸化,明显减少Bax、活化caspase-3、活化caspase-9在脊髓神经元中的表达,降低TUNEL阳性细胞的比例,增加Bcl-2的水平,有效改善SCI后的运动功能恢复。Tural等[21]报道,臭氧治疗SCI可以增加β-catenin表达,并减少神经损伤和白质空洞,促进运动功能的恢复。Shen[22]等认为褪黑素激活Wnt/β-catenin信号通路减轻SCI后神经凋亡。SCI后内源性神经干细胞向神经元分化可以弥补一部分凋亡的神经元,降钙素基因相关肽(calcitonin gene-related peptide,CGPR)促进大鼠脂肪源性干细胞(adipose-derived stem cells,ADSCs)向神经细胞分化,Wnt-3a,Wnt-5a和β-catenin通过上调CGPR的表达介导ADSCs神经分化的过程[23]。
2.3 Wnt信号通路参与SCI后的轴突再生和胶质瘢痕形成过程
OPCs的激活是SCI后胶质瘢痕形成的重要原因。条件性敲除OPCs的β-catenin基因可抑制Wnt信号通路,减少OPCs的反应和小胶质细胞的激活,减少胶质瘢痕形成,形成一个支持轴突生长的环境[14]。大鼠脊髓损伤和斑马鱼脊髓横断后β-catenin在损伤区域的表达增加,促进放射状胶质细胞的神经分化、轴突再生[24-25]。Dickkopf-1(DKK-1)过表达会抑制 Wnt/β-catenin信号通路,抑制SCI后运动功能的恢复、轴突再生和胶质桥的形成[25]。Ryk是一个轴突引导相关的Wnt受体[26-27]。Gonzá lez等[28]报道,SCI可以诱导Ryk受体在损伤区域内活化的星形胶质细胞、小胶质细胞、巨噬细胞,NG2+神经胶质前体细胞、纤连蛋白细胞、少突胶质细胞和轴突中表达。通过注射Ryk的单克隆抗体或者敲除Ryk基因,增加了皮质脊髓轴突附属分支的前突触突变点,促进前肢运动功能的恢复。抑制Ryk受体同时结合康复训练,更有利皮质脊髓束的轴突再塑造和皮质回路的再形成[29]。Liu等[30]报道,Ryk受体在损伤区域内的皮质脊髓束轴突中表达,注射Ryk功能抑制抗体明显促进损伤区域皮质脊髓束侧支的发芽。
2.4 Wnt信号通路参与脊髓损伤后的骨质丢失过程
SCI后机械荷载消失、神经损伤、肌肉瘫痪和激素水平改变造成严重的骨丢失,小梁连接和板状小梁结构减少,导致骨质疏松。这一过程与SCI后Wnt信号通路受到抑制有密切关联[31]。据报道,SCI后成骨细胞中Wnt3a、LRP5、FZD5、Tcf7mRNA水平降低,Wnt信号抑制剂Dkk-1的表达明显增加,Dkk-1的浓度与局部骨质丢失呈正相关[31]。血清骨硬化蛋白(Sclerostin,SCL)是骨细胞产生的Wnt信号通路拮抗剂。血清骨硬化蛋白抗体(Sclerostin antibody,SCL-Ab)可以阻止SCI诱导的骨丢失,同时增加骨质的形成和骨细胞的表面积,保留骨细胞的形态学和结构[32-33]。SCL-Ab抑制破骨细胞,促进成骨细胞中LRP5、破骨细胞抑制因子(osteoprotegerin,OPG)的mRNA水平,促进OPG/RANKL比值增加,减少Dkk-1的mRNA水平[33]。Yang等[34]报道激活Wnt/β-catenin信号通路可缓解局部骨质丢失。
3 姜黄素调控Wnt信号通路在SCI后的作用
姜黄素是一种从姜黄属中药如姜黄、莪术和郁金等植物中提取的橙黄色天然植物多酚。姜黄素具有无毒、抗炎、抗氧化、抗肿瘤和降脂等优点,能与许多靶点结合产生多种生物学效应,缺点是水溶性差,生物利用度低等[35]。
3.1 姜黄素与Wnt信号通路
Wnt配体作为Wnt/β-catenin信号通路的起点,姜黄素可以通过直接激活Wnt配体,促进Wnt配体在细胞外的浓度升高,进而激活Wnt/β-catenin信号通路。姜黄素可以通过激活丝氨酸/苏氨酸激酶(serine/threonine kinase,Akt)启动GSK-3β的磷酸化,从而抑制βcatenin降解复合物,达到激活Wnt信号通路的作用[36]。姜黄素可以明显提高Wnt3a和β-catenin的蛋白质和RNA浓度,促进神经干细胞向神经元的分化[37]。参见图1。
3.2 姜黄素调控Wnt信号通路治疗脊髓损伤
姜黄素是Wnt的一个天然调节剂,通过激活Wnt信号抑制SCI后的炎症反应,达到治疗效果。姜黄素通过抑制iNOS,COS-2和NF-κB的激活,抑制NO、TNF-α、IL-6的炎症因子的释放,激活Nrf2,明显减少NF-κB的活性,减少炎症因子的产生,减少运动障碍的严重程度,减少脊髓水肿和凋亡[38]。β-catenin对NF-κB诱导的转录活动起抑制作用[39]。Ni等[40]报道,姜黄素治疗可明显下调SCI诱导的TLR4/NF-κB炎症信号通路相关因子,并且明显改善后肢运动功能障碍、脊髓水肿与凋亡。姜黄素通过激活Wnt3a以及 Akt,使P13k/Akt信号通路和Wnt/β-catenin信号通路产生交叉,诱导GSK-3β磷酸化,促进β-catenin在胞质内的累积,激活TCF/LEF的转录活动,起到保护神经细胞的效果[37]。Tiwari等[41]认为,负载姜黄素纳米颗粒可以通过激活Wnt/β-catenin信号通路诱导神经干细胞的增殖和神经元的分化,增加β-catenin活性和核转录活动。同时,姜黄素可以促进SCI后大鼠后肢功能恢复,促进Tuj1阳性神经元数量增加,减少损伤区域星形胶质细胞胶质纤维酸性蛋白(glial fibrillary acidic protein,GFAP)和硫酸软骨素蛋白多糖(chondroitin sulfate proteoglycans ,CSPG)的表达,促进Wnt3a和β-catenin的表达量,通过激活Wnt/β-catenin信号通路促进内源性神经干细胞向神经元分化,减少胶质瘢痕的形成,促进脊髓损伤的修复[42]。
脊髓损伤后,破骨细胞活性和数量的增加,骨吸收和骨形成的平衡被打破,造成严重的骨质丢失,增加病人骨折的风险。Wnt抑制剂如Dkk-1、SCL等的表达增加,抑制Wnt/β-catenin信号通路,影响成骨细胞和骨细胞分化,造成骨质丢失[31]。姜黄素通过激活Wnt/βcatenin信号通路抑制SCI后的骨质丢失,增加远端股骨中Wnt3a、Lrp5和ctnnb1的mRNA水平,上调β-catenin的蛋白表达,缓解SCI后大鼠局部骨质丢失[43]。Yamaguchi等[44]报道,姜黄素类似物UBS109通过刺激成骨细胞钙化和抑制破骨细胞形成来抑制骨质丢失。Elburki[45]等报道,姜黄素可以抑制炎症,减少NF-κB和p38 MAPK,抑制骨丢失。
综上所述,姜黄素可以在SCI中调控Wnt信号通路,减轻炎症反应、抑制细胞凋亡、抑制骨质丢失,促进脊髓损伤后的运动功能恢复。
4 问题与展望
Wnt信号通路是SCI治疗的一个重要调节点,姜黄素通过刺激Wnt配体、Akt和抑制GSK-3β来激活Wnt信号通路,使β-catenin在胞内累积,促进目标基因表达,起到抑制SCI后炎症反应、细胞凋亡、骨质流失和促进轴突再生的效果。姜黄素由于其水溶性差、生物利用度低、药代动力学差等问题,应用于临床疾病治疗尚需进一步改进和优化。开发出水溶性好、生物活性和利用度高的姜黄素衍生物,进一步明确姜黄素在SCI后对信号通路的调控机制,对于SCI治疗具有广阔的应用前景。
猜你喜欢
浙江临床医学(2021年12期)2021-11-29 14:43:18
神经损伤与功能重建(2020年11期)2020-12-01 05:01:54
Medical Data Mining(2019年2期)2019-07-16 04:36:08
中成药(2018年2期)2018-05-09 07:19:43
中成药(2018年3期)2018-05-07 13:34:37
湖南中医药大学学报(2016年1期)2016-12-01 04:08:21
磁共振成像(2015年1期)2015-12-23 08:52:21
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:27
西南军医(2015年1期)2015-01-22 09:08:25
中华神经创伤外科电子杂志(2015年1期)2015-01-21 09:09:24
