
运动激活自噬对小鼠骨骼肌抗氧化防御功能的影响
2018-07-18 09:23:28王平ChunGuangLI崔迪邱守涛漆正堂李娅晖丁树哲
中国运动医学杂志 2018年1期
王平Chun-Guang LI崔迪 邱守涛 漆正堂 李娅晖 丁树哲
1杭州师范大学体育与健康学院(杭州 311121)
2 National Institute of Complementary Medicine,Western Sydney University
3华东师范大学青少年健康评价与运动干预教育部重点实验室
4上海交通大学生命科学学院
骨骼肌占机体总重的40%~50%,是一种高度可塑性组织,运动刺激可使肌肉发生形态学和生理生化功能方面的适应性显型变化[1]。已有研究报道,耐力运动可明显增加老年大鼠腓肠肌总超氧化物歧化酶(TSOD)和铜锌SOD(CuznSOD)的活性,同时增强腓肠肌线粒体锰SOD(MnSOD)的活性[2]。耐力运动明显增加3月龄大鼠股外侧深肌、股外侧浅肌和跖肌MnSOD的活性和股外侧浅肌的CuznSOD活性[3]。一次性力竭运动明显增加大鼠股外侧深肌MnSOD mRNA的表达,同时增加股外侧浅肌MnSOD蛋白含量,股外侧深肌和股外侧浅肌CuznSOD蛋白含量[4]。故运动使骨骼肌抗氧化防御功能的积极适应是维持骨骼肌健康乃至整个机体健康的重要前提和基础,但运动性骨骼肌有益适应的潜在分子机制还不是很清楚。
有研究发现,自噬参与骨骼肌氧化损伤等应激反应[5]。Dobrowolny等[6]为了证实自噬与骨骼肌抗氧化防御功能之间的关系,建立了CuznSOD基因突变小鼠模型,结果证实自噬与抗氧化防御功能之间关系密切。运动可适度激活自噬,它可将骨骼肌细胞内部分细胞器、蛋白质形成分隔膜包绕,形成自噬体,然后通过细胞骨架微管系统运输至溶酶体消化降解,并有效清除骨骼肌收缩过程中产生的有害的活性氧,从而维持骨骼肌细胞功能稳态[7]。
目前而言,运动性自噬在骨骼肌氧化应激过程中是否必须,运动、自噬和骨骼肌抗氧化防御功能之间潜在的分子机制还不是很清楚。本研究建立一次性力竭运动小鼠模型,通过检测骨骼肌自噬相关因子Beclin1、P62、Bcl2的表达变化和T-SOD、CuznSOD、MnSOD活性和T-AOC的含量变化,及P62的蛋白表达与骨骼肌TAOC含量和CuznSOD活性之间的相关性分析,探讨运动能否激活自噬和运动性自噬与骨骼肌抗氧化防御功能之间发生交互作用的控制节点。预期成果将进一步阐明运动性自噬是维持骨骼肌氧化应激稳态的一条重要调控路径,增进对骨骼肌显型适应调控机制的进一步认识,还有可能为巩固和优化骨骼肌质量及其功能的分子机制提供新思路。
1 材料与方法
1.1 实验动物与分组
4周龄清洁级健康雄性ICR(Institute for Cancer Research)小鼠30只,体重25.17± 2.46 g,购自上海斯莱克动物实验中心,生产许可证号为:SCXK(沪)2007-0005,使用许可证:SYXK(沪)2004-0001。小鼠常规分笼(5只/笼),国家标准啮齿类动物常规饲料(上海斯莱克动物实验中心提供)饲养、自由饮水、饮食,室温为20℃~23℃,相对湿度为50%~70%,光照12 h/天,更换垫料2~3次/周,保持通风。ICR小鼠专用饲料和垫料均由上海生工生物技术有限公司提供。小鼠适应性喂养3天后,随机分为安静对照组(C组,n=6)和运动组(n=24),运动组小鼠进行一次性力竭跑台运动,运动力竭后再次分为运动后即刻组0 h(n=6)、6 h组(n=6)、12 h组(n=6)和24 h组(n=6)。
1.2 运动方案
运动组小鼠进行适应性跑台训练3天,坡度为0℃,速度为15 m/min,持续30 min/day。正式运动参照Bedford根据鼠体重/摄氧量回归方程所建立的递增运动负荷训练方案[8],按以下程序运动:第1级负荷:0°,8.2 m/min(相当于53%VO2max),15 min;第2级负荷:5°,15 m/min(相当于64%VO2max),15min;第 3级负荷:10°,19.3 m/min(相当于 76%VO2max)运动至力竭。运动时使用毛刷刺激,使动物保持在跑道前1/3处,以保证运动强度。待小鼠持续运动至不能坚持原跑速,刺激驱赶无效,停跑后体征表现为俯卧位,呼吸急促,神情倦怠,对刺激反应迟钝时,判定为力竭[9]。整个实验过程中每只小鼠跑的力竭时间被记录,平均力竭时间为173.83±13.75 min。
1.3 取材
安静对照组在安静状态、运动组于运动后对应时间点颈椎脱臼处死小鼠,迅速取左侧腓肠肌称重,用于生化测试,右侧腓肠肌切分若干份装入已标记好的冻存管,迅速置于液氮中,然后转到-80℃冰箱保存,用于检测转录基因表达和蛋白表达。
1.4 生化测定
腓肠肌T-SOD、CuznSOD和MnSOD活性均采用黄嘌呤氧化酶法测定,T-AOC含量采用比色法测定,生化具体操作均严格按照说明书进行,均采用Tecan 2000型酶标仪检测,蛋白定量采用BCA法。
1.5 Real-Time PCRe PCR检测
冰上取腓肠肌约40 mg采用Invitrogen Trizol法提取总RNA,总RNA的OD260/OD280的比值采用紫外分光光度计检测并计算选取符合Real-time PCR要求的RNA进行逆转录;逆转录时取5 μl RNA样品,配置10 μl的总反应体系,使用TOYOBO FSQ101反转录试剂盒合成第一链cDNA(15℃,5 min;37℃,15 min;85℃,5 min);以合成的cDNA为模板,以GAPDH为内参,在ABI StepOne型实时荧光定量PCR进行荧光定量,反应条件为:95℃15 s,61℃30 s,72℃45 s,40个PCR循环。PCR仪给出各反应孔的Ct值,以GAPDH为内参,根据公式2-ΔΔCt计算各样品目的基因的相对表达量,其中ΔΔCt=(Ct实验组目的基因-Ct实验组内参基因)-(Ct对照组组目的基因-Ct对照组内参基因)。荧光染料为TOYOBO QPK201 SYBR GREEN,实验所用引物均参照GeneBank数据库,由上海生物工程有限公司设计并合成。
1.6 Western Blot Blot检测
取腓肠肌约50 mg冰上剪碎入研磨管中,按照Beyotime P0013 Western及IP裂解液说明书加入0.5 ml裂解液(含1mM PMSF),OMINI Bead Ruptor 24 型磁珠匀浆机匀浆,14000 g离心15 min,上清转入离心管,BCA法测定蛋白浓度后蛋白变性。采用10%、12%分离胶电泳,湿转法将蛋白转至PVDF膜;使用5%脱脂奶粉封闭,一抗(Anti-Beclin1、Anti-Bcl2和Anti-P62稀释比例分别为1∶1000、1∶200和1∶200,购于美国CST或Santa Cruz公司)4℃孵育12 h,TBST缓冲液清洗后,HRP标记二抗室温孵育2 h,Millipore ECL超敏试剂盒显影,AlphaFC2型凝胶成像系统进行冷光扫膜,使用FluorChem FC2软件对所捕捉图像进行灰度值分析,内参为GAPDH,实验所用抗体购于Santa Cruz或CST公司。

表1 各基因引物序列
1.7 数据统计
各组检测数据录入Excel 2013,每组数据结果用均数±标准差(±s)表示,用GraphPad Prism5软件进行数理统计和图生成,组间比较采用单因素方差分析,以P<0.05为显著性水平,P<0.01为极显著性水平。
2 结果
2.1 运动对小鼠腓肠肌自噬相关因子Beclin1、Bcl2和P62 mRNA表达的影响
与安静对照组相比,运动后0 h、6 h小鼠腓肠肌Beclin1 mRNA表达量均出现极显著升高(P<0.01),12 h、24 h均未出现显著升高(P>0.05,图1A);Bcl2 mRNA在运动后0 h、6 h、12 h和24 h的表达量均未出现显著变化(P>0.05,图1B);P62 mRNA的表达量在运动后24 h出现极显著升高(P<0.01),0 h、6 h和12 h均未出现显著升高(P>0.05,图1C)。
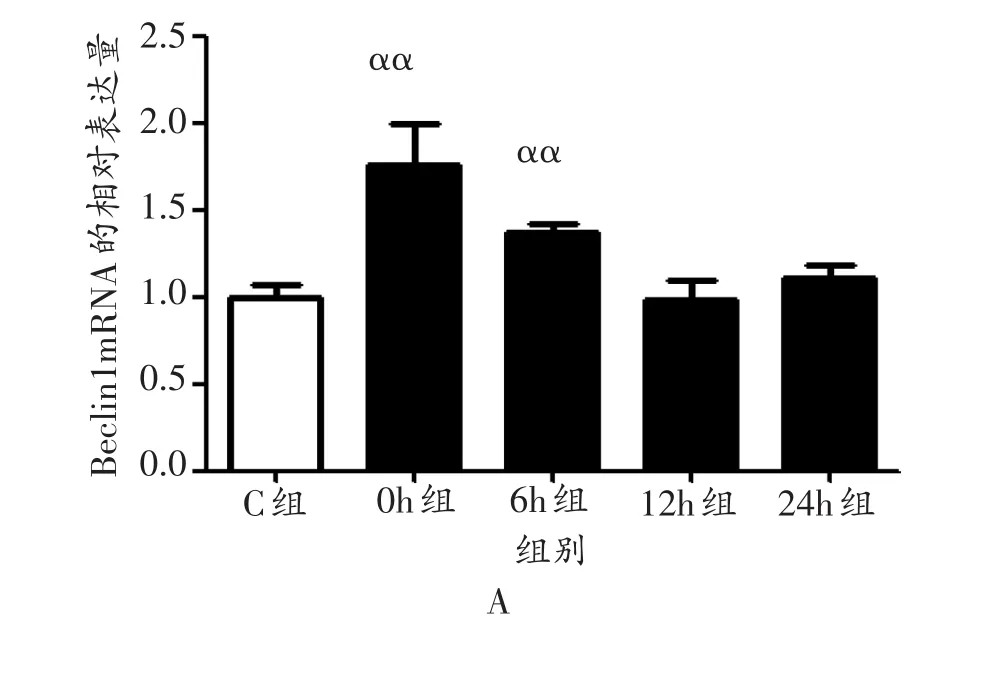
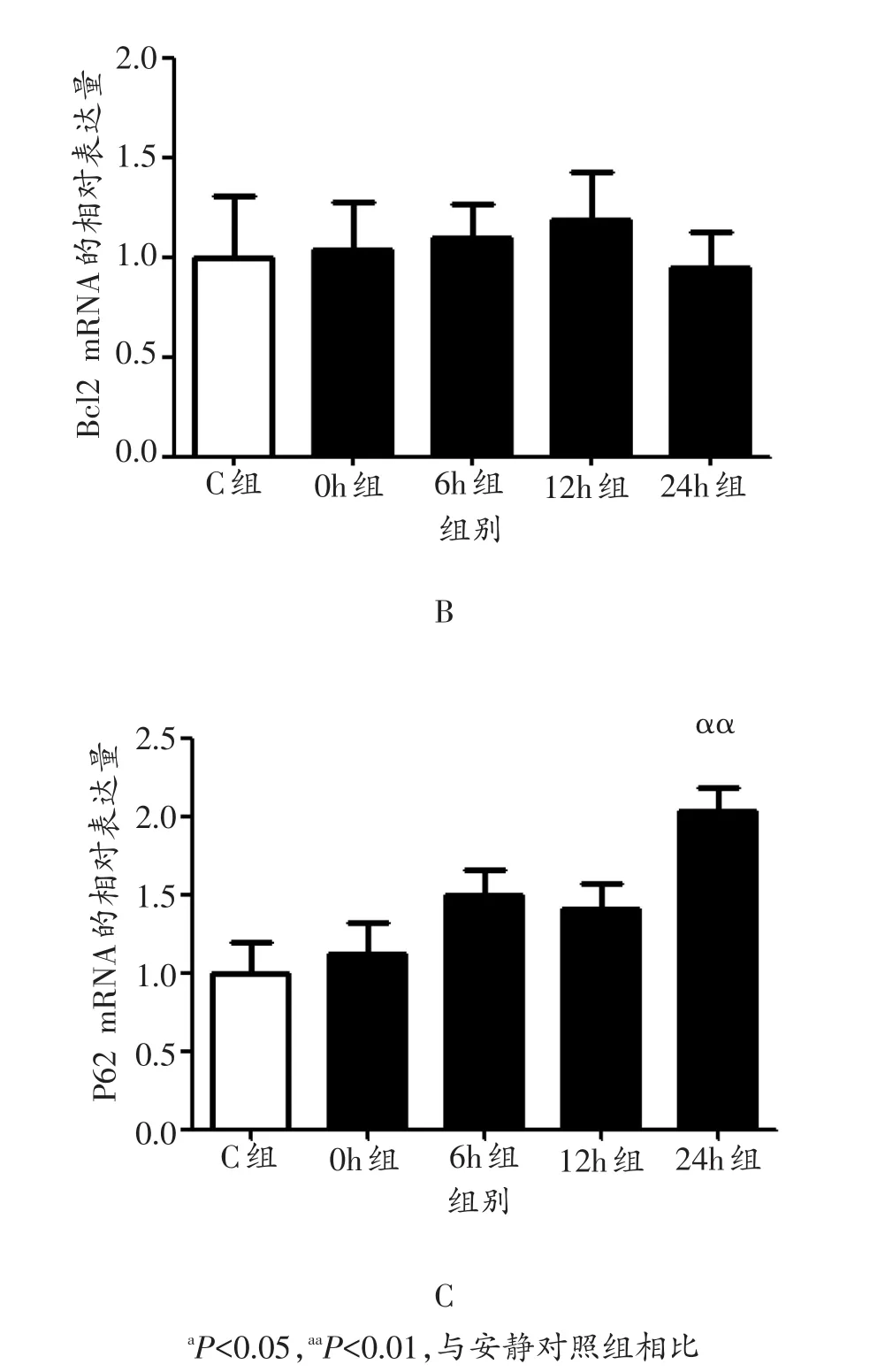
图1 各组小鼠腓肠肌Beclin1 mRNA、Bcl2 mRNA和P62 mRNA表达量
2.2 运动对小鼠腓肠肌自噬相关因子Beclin1、Bcl2和P62蛋白表达的影响
与安静对照组相比,小鼠腓肠肌Beclin1蛋白表达在运动后0 h、6 h、12 h和24 h均未出现显著变化(P>0.05,图2A);Bcl2蛋白表达在运动后0 h、6 h和12 h出现显著升高(P<0.05或P<0.01),24 h未出现显著变化(P>0.05,图2B);P62蛋白表达在运动后0 h、12 h和24 h均出现显著下降(P<0.05或P<0.01),6 h未出现显著变化(P>0.05,图2C)。
2.3 运动对小鼠腓肠肌T-SOD、CuznSOD、MnSOD活性和T-AOC含量的影响
与安静对照组相比,小鼠腓肠肌T-SOD活性在运动后6 h出现极显著下降(P<0.01),0 h、12 h和24 h均未出现显著变化(P>0.05,图3A);CuznSOD活性在运动后0 h显著增加(P<0.05),6 h显著下降(P<0.05),12 h和24 h均未出现显著变化(P>0.05,图3B);MnSOD活性在运动后0 h出现极显著升高(P<0.01),6 h、12 h和24 h均未出现显著变化(P>0.05,图3C);T-AOC含量在运动后6 h、12 h和24 h均出现显著升高(P<0.05或P<0.01),0 h未出现显著变化(P>0.05,图3D)。
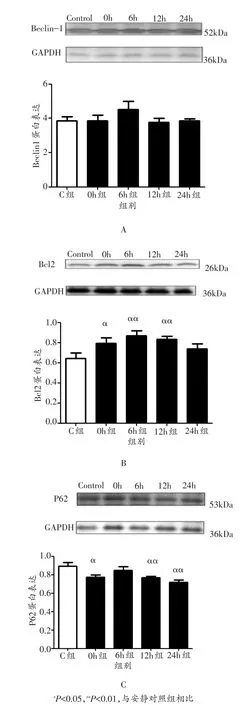
图2 各组小鼠骨骼肌Beclin1、Bcl2和P62蛋白表达变化
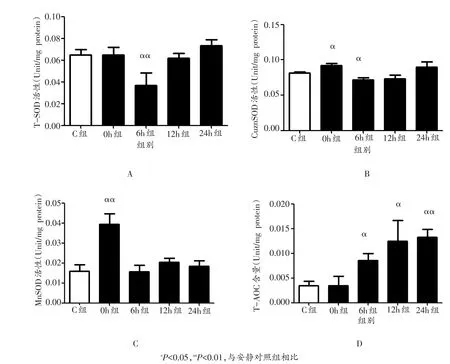
图3 各组小鼠骨骼肌T-SOD、CuznSOD和MnSOD的活性及T-AOC含量的变化
2.4 自噬相关因子P62蛋白表达与小鼠腓肠肌T-AOC含量和CuZnSOD活性的相关性分析
小鼠腓肠肌P62蛋白表达与T-AOC含量之间呈负相关(r=-0.6239,P<0.01,图 4A);P62蛋白表达与CuZnSOD活性之间虽然未达到统计学差异(r=-0.3379,P=0.0678>0.05,图4B),但具有相关性趋势。
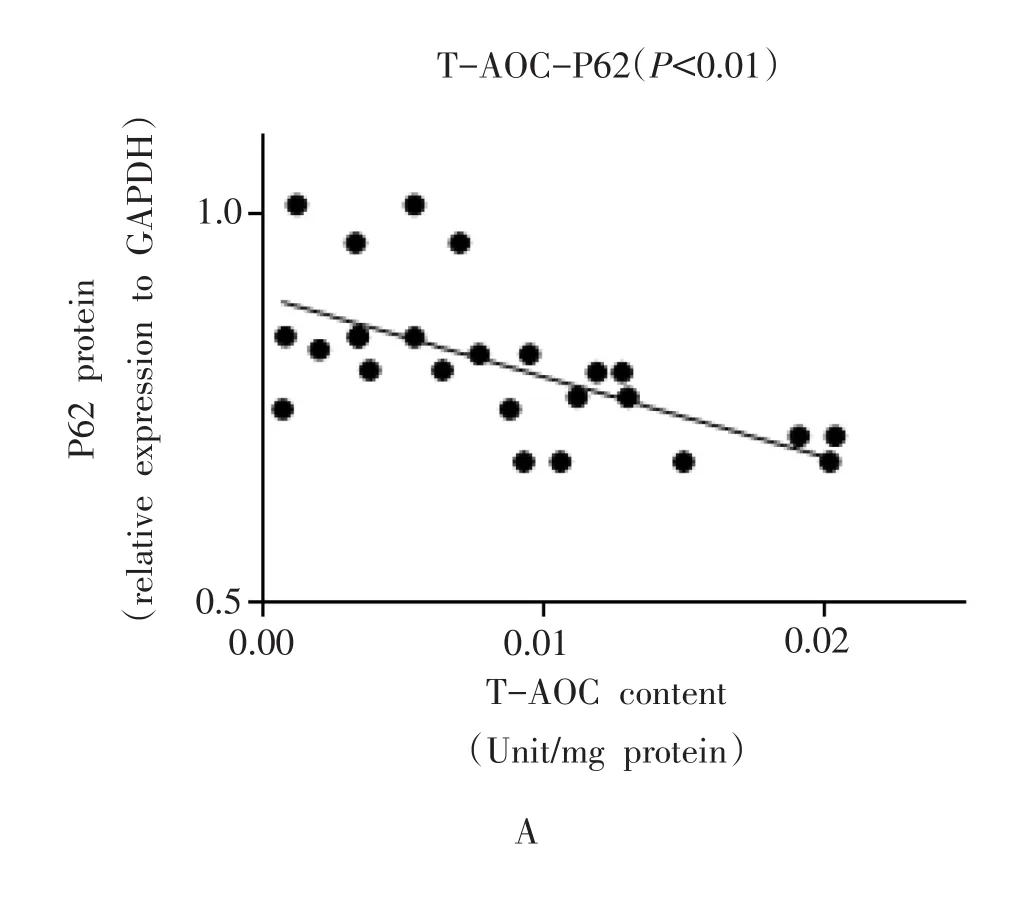
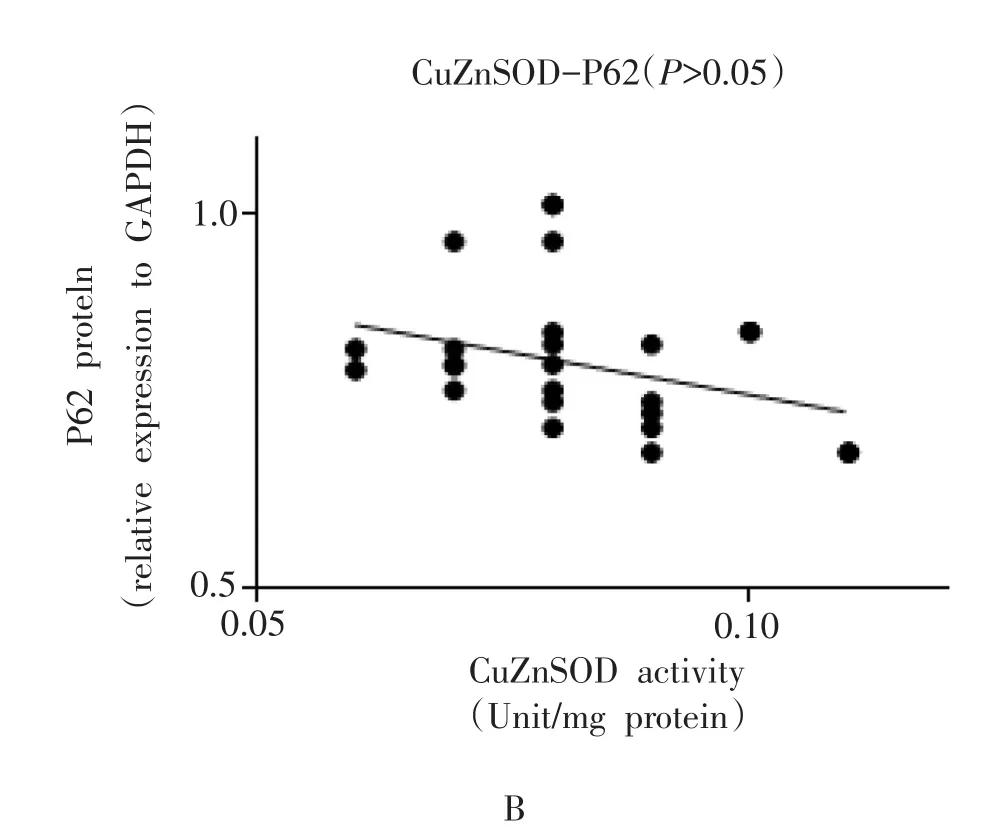
图4 小鼠腓肠肌T-AOC含量、CuZnSOD活性与P62蛋白表达之间的相关性分析
3 分析与讨论
3.1 运动对骨骼肌自噬相关因子Beclin1和P62表达的影响
自噬在骨骼肌肌纤维分解代谢中的作用引起学者们的普遍关注,关于骨骼肌分解代谢模型(饥饿、缺氧和恶液质)的研究发现,自噬激活同时伴随着蛋白转换增加[10]。适宜负荷的运动可提高骨骼肌细胞自噬水平,分析其机制发现,自噬除了降解运动过程中产生和积累的破损、衰老的细胞器,还和合成或折叠错误的蛋白质有关,从而为肌纤维更新代谢提供一定的能量与合成所需的底物,同时抑制骨骼肌细胞凋亡和死亡[11]。但关于骨骼肌运动适应过程中自噬调控的分子机制还不是很清楚。Beclin1(又称Atg6)在真核细胞中高度保守,属于自噬相关蛋白家族,在自噬体(autophagosome)的形成过程中是必需的。它可介导其它自噬蛋白定位于吞噬泡(phagophore),调控哺乳动物自噬体的形成与成熟。Beclin1表达水平上升提示自噬活性增加[12]。而P62位于自噬体,其作用是募集蛋白质进入自噬体进行降解,P62水平下降提示自噬活性增加,而P62积累提示自噬缺陷[13]。Jeong等[14]研究发现,耐力游泳运动明显增加小鼠骨骼肌Beclin1蛋白表达,明显降低P62蛋白表达。Hecongcong[15]研究发现,一次性力竭跑台运动明显降低小鼠骨骼肌P62蛋白表达。Vitor等[16]研究发现,耐力转轮运动显著增加小鼠骨骼肌Beclin1蛋白表达,并显著降低P62蛋白表达,提示运动可有效增加骨骼肌自噬水平以维持骨骼肌收缩活动和新陈代谢所需。
本研究发现,一次性力竭跑台运动后0 h和6 h小鼠骨骼肌Beclin1 mRNA转录水平显著提高,0 h、12 h和24 h P62蛋白表达显著降低,提示运动引起的Beclin1 mRNA表达增加反应早,但延续时间相对短,而P62蛋白表达下调反应早且延续的时间相对长。因此Beclin1 mRNA和P62蛋白表达可以同时作为一次性力竭运动后早期自噬反应的敏感性指标;P62蛋白表达还可以作为一次性力竭运动后延迟性自噬反应的敏感性指标。这些结果提示不同的自噬指标对一次性力竭运动后的不同时程敏感性是不同的,故选择合适地能够反映自噬的指标非常重要。本实验中Beclin1 mRNA和P62蛋白表达的变化提示一次性力竭跑台运动可激活骨骼肌自噬,是骨骼肌自噬强有力的诱导剂,这与先前的研究结果报道一致。但文献中也有相反的报道,Kim等[17]研究发现,一次性力竭运动显著降低小鼠骨骼肌Beclin1蛋白表达,提示一次性力竭运动也可能减弱骨骼肌自噬信号途径。造成上述研究结果不一致的原因可能如下:第一,运动过程中能量消耗的不同(不同实验中的运动强度和运动持续时间均不同)可能决定自噬的活性。腺苷酸活化蛋白激酶(AMP activated protein kinase,AMPK)是能量状态的敏感感受器,也是细胞自噬的重要激活因子。如果运动使骨骼肌肌纤维内AMP/ATP比值显著增加,则可激活其下游信号分子AMPK,后者可通过磷酸化Raptor的Ser722/Ser792位点或磷酸化TSC2 Thr1227/Ser1345位点抑制mTOR,诱导细胞自噬。但如果运动时能量消耗是暂时的且储存在细胞中的能源物质没有耗尽时,自噬有可能不被激活,甚至降低[18]。故不同能量消耗可引起不同的自噬反应。第二,运动前肌糖原含量不同,自噬反应可能也不同。已有研究发现,运动前肌糖原含量与AMPK活性呈负相关[19]。第三,自噬变化可能与不同肌纤维类型、处死时间、样本的收集、饮食和年龄等内在和外在因素有关系[20]。
本研究发现一次性力竭运动后24 h骨骼肌P62 mRNA表达增加,与Tam等[21]的研究结果一致,但蛋白表达明显下降。分析其原因可能是在自噬的降解阶段,P62基因在转录延长阶段消耗了一部分(降解增加),其目的是为了避免自噬相关因子耗尽[22];也有可能P62的作用除了作为自噬适应物之外,还有其他的生物学作用,包括在Ras/Raf/MAPK和NF-κB通路、氧化应激和肿瘤生成等信号转导通路中发挥作用[23]。
本研究还发现,一次性力竭运动并没有使骨骼肌Beclin1蛋白表达发生显著变化,与Jamart[24]的研究结果一致。后者研究发现,禁食并没有引起Beclin1蛋白表达的变化。同样,Tam[21]发现禁食后的耐力运动并没有导致骨骼肌Beclin1蛋白表达发生变化。运动性骨骼肌自噬的变化可能与禁食时诱导的自噬一致。分析Beclin1 mRNA表达增加而蛋白表达不增加的原因可能如下:其一,蛋白表达水平的变化与转录及转录后水平调控、翻译及翻译后水平调控及蛋白质降解等均有关系;其二,蛋白的表达不一定与它相对应的mRNA水平呈现同步表达[25]。已有人对同一基因的蛋白表达与mRNA表达是否一致做了相关性研究,结果发现,相关系数为0.46~0.68[26],但其具体分子机制还需要进一步研究。
3.2 运动激活自噬相关因子Bcl2和P62对骨骼肌抗氧化防御功能的影响
B细胞淋巴瘤/白血病-2(Bcl2,B cell lymphoma/leukemia-2)是一种含有多个BH结构域的抗凋亡蛋白。有研究发现,它在调控细胞自噬过程中具有重要作用,通过与Beclin1的BH3结构域结合形成复合物,抑制Beclin1诱导细胞自噬。在哺乳动物细胞中,Bcl2与Beclin1的分离对于自噬的诱导至关重要[27]。Hecongcong[15]使用免疫沉淀反应研究发现运动15分钟Bcl2与Beclin1复合物明显下降,运动30分钟,其复合物基本检测不到。
有研究发现,Bcl2还可通过多种机制调控细胞抗氧化防御系统[28]。Veis[29]的研究发现,Bcl2基因敲除小鼠的抗氧化防御功能存在缺陷。Kane等[30]研究发现,Bcl2过表达提高细胞中谷胱甘肽(GSH)含量。Merad[31]采用过表达技术上调细胞中Bcl2蛋白的表达,以模拟SOD过表达的效应,结果发现,Bcl2与SOD具有正相关关系。本实验研究发现,一次性力竭运动后0 h、6 h和12 h骨骼肌Bcl2蛋白表达明显增加,提示运动引起的Bcl2蛋白表达增加反应早且延续时间相对长,它也可作为一次性力竭运动后即刻且延迟到运动后12 h自噬反应的敏感性指标。同时0 h骨骼肌MnSOD、CuznSOD活性均显著增加,6 h、12 h和24 h骨骼肌TAOC含量也显著增加,Bcl2蛋白表达的增加趋势基本与骨骼肌抗氧化防御功能增加的趋势一致,上述研究结果提示:当主要位于线粒体膜的Bcl2蛋白表达增加时,MnSOD和CuznSOD活性增加,骨骼肌总T-AOC含量也明显增加,这样可以降低细胞内活性氧(ROS)的水平,阻止氧化应激的细胞损伤[28]。Bcl-2还能够与GSH结合,清除线粒体氧化呼吸链上漏出的超氧阴离子等ROS[32]。有研究发现Bcl-2能够影响细胞色素C氧化酶Ⅳ(COXⅣ)的活性进而影响电子呼吸链的功能及线粒体超氧阴离子的水平[28],同时,本研究还发现,P62蛋白表达与骨骼肌T-AOC含量呈相关性,提示激活自噬相关因子Bcl2和P62的蛋白表达与骨骼肌抗氧化防御功能有关,其过程可能与Bcl2和P62的蛋白表达的、时间点活化有关,因此,Bcl2和P62或共同调控和增强骨骼肌的抗氧化防御功能,然而具体生物学分子机制仍需要进一步研究。
本实验研究结果还发现,一次性力竭运动并没有引起骨骼肌Bcl2 mRNA表达变化,但Bcl2蛋白表达增加,分析原因:其一,可能与mRNA的产生效率有关。通常情况下,mRNA的产生效率远远低于蛋白质,某一给定的mRNA每小时只能复制2次,但同样时间可以拷贝数十个相应的蛋白;其二,mRNA稳定性差,mRNA半衰期通常是2.6~7 h,而蛋白是46 h[33]。也有不同的报道,发现一次性力竭游泳运动使小鼠骨骼肌Bcl2 mRNA表达下降,可能是由于不同的运动方式、不同的运动强度与运动时间导致小鼠骨骼肌Bcl2转录基因表达产生不同的反应[34]。
综上所述,Bcl2和P62在自噬过程中可能起着关键的调节作用,在骨骼肌抗氧化防御功能中也有重要作用,提示自噬和抗氧化防御系统之间存在着复杂的联系。关于运动、Bcl2、P62和抗氧化防御功能的确切分子机制,还需要进一步研究。
4 结论
一次性力竭跑台运动可明显激活骨骼肌自噬相关因子Beclin1 mRNA和P62蛋白的表达,可适度提高自噬,但不同自噬指标对运动的敏感时程性不同,且激活后活性延迟的时间不同;一次性力竭运动可增强骨骼肌抗氧化防御功能,骨骼肌抗氧化防御功能的增加与自噬相关因子P62蛋白的表达具有相关性。
猜你喜欢
分子诊断与治疗杂志(2023年8期)2023-09-07 12:00:34
中成药(2018年10期)2018-10-26 03:41:40
中成药(2018年3期)2018-05-07 13:34:34
岁月(2017年7期)2017-07-18 18:52:11
中国老年学杂志(2017年10期)2017-06-15 19:07:40
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
磁共振成像(2015年5期)2015-12-23 08:52:53
医学研究杂志(2015年5期)2015-06-10 06:43:26
