
适度运动增强学习记忆研究进展
2018-07-18 09:23:34王浩权韩静
中国运动医学杂志 2018年1期
王浩权 韩静
陕西师范大学现代教学技术教育部重点实验室(西安 710000)
学习记忆作为大脑最重要的高级认知功能之一。如何改善和提高学习记忆能力是心理学、神经生物学和医疗保健等领域研究者们共同关注的问题。近些年来,大量的动物和人类的研究都表明,长期适度运动对学习记忆有积极作用[1,2],而运动对学习记忆的增强涉及外周和中枢系统,其分子机制相当复杂,至今尚无较为系统和全面的结论。突触作为学习记忆形成的结构基础,一直以来都是相关研究的焦点,也是运动增强学习记忆的内在机制研究主要关注的脑内结构,随着相关动物研究的不断开展,运动增强学习记忆以突触为结构基础的分子机制也逐渐清晰。研究发现脑源性营养因子(brain derived neurotrophic factor,BDNF)、胰岛素样生长因子-1(insulin-like growth factor-1,IGF-1)和血管内皮生长因子(vascular endothelial derived growth factor,VEGF)三种调节因子在运动增强学习记忆的过程中起到十分关键的作用。本文以这三种调节因子为主线,梳理运动增强学习记忆以突触为结构基础的分子机制,并就动物与人类相关研究在方法和结果的异同进行比较,分析开展人类相关研究所遇到的瓶颈及进一步研究可能的方向。
1 运动增强学习记忆相关的三种重要的调节因子
1.1 BDNF
BDNF是一种十分重要的神经营养因子(neurotrophin,NT),由神经所支配的组织和星形胶质细胞产生,且为神经元生长与存活所必需的蛋白质分子,对支持神经元功能完整性有重要作用[3]。另外,BDNF有高度保守的基因和功能,在突触形成和大脑认知活动中起到非常重要的作用[4]。研究者通过研究不同的大鼠运动模型发现,运动可以提高大鼠海马BDNF的表达量和学习记忆能力[5,6]。进一步研究发现,BDNF可以与TrkB受体结合,通过丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和磷脂酰肌醇 3激酶(phosphatidylinositol 3 kinase,PI3K)信号转导途径促进生长因子、离子通道等的表达,激活突触后膜α-氨基-3羟基-5甲基-4异恶唑受体(α-amino-3-hydroxy-5-methylisoxazole4-propionate receptor,AMPAR)和N-甲基-D-天冬氨酸受体(N-methyl-D-aspartic acid receptor,NMDAR),提高突触可塑性,促进突触的形成[7]。除此之外,BDNF还可以提高突触蛋白-I(synapsin-I)和突触素(synatophysin)的表达量,增加突触前膜递质的释放,进而影响学习记忆[8,9]。而对于运动具体如何调节BDNF的表达量,有研究表明运动可以通过IGF-1提高BDNF的表达量[10],还有研究认为运动可以通过组织类纤维酶原激活物(tissue type plasminogen activator,tPA)增加脑源性营养因子前体(pro-Brain derived neurotrophic factor,proBDNF)向 BDNF转化,提高BDNF的量[5]。而proBDNF与BDNF在介导运动对学习记忆的影响中有相互拮抗的作用[11]。因此,tPA介导的proBDNF与BDNF之间的转化或许是BDNF介导的运动增强学习记忆的关键。另外,有研究发现运动后骨骼肌BDNF的表达量也会显著增加[12],这提示我们参与运动增强学习记忆的BDNF也可能来自于外周系统。因此,进一步探明运动与tPA之间的关系,明确参与运动增强学习记忆过程中的BDNF的准确来源,对于更加全面地了解运动增强学习记忆的分子机制非常重要。
1.2 IGF-1
IGF-1是一种多功能细胞增殖调控因子,在中枢和外周系统中均有表达,但IGF-1在中枢系统中的表达量会随着个体的成熟而减少,机体中70%以上的IGF-1来自肝脏和肌肉等组织,其在细胞的分化、增殖及个体的生长发育中具有重要作用。另外,IGF-1还可以通过血脑屏障,参与脑内微血管结构和功能调节,并对神经元之间的活动及其可塑性有重要的调节作用[13,14]。近些年来的研究表明,运动可以提高循环系统中IGF-1的量,同时提高海马区神经发生及BDNF的表达量,并诱导c-fos基因表达,增强学习记忆能力,而封闭循环系统IGF-1对中枢系统的作用,运动对海马区神经发生的提高及学习记忆的增强都同样被消除,目前虽然不能完全排除运动后中枢系统自身IGF-1表达量的提高,但可以确定外周源的IGF-1在运动增强学习记忆的过程中起到了主要作用[10,15]。进一步研究发现运动可以同时提高血液及海马区IGF-1的量,但肝脏IGF-1的分泌量并没有受到运动的影响[16],这提示我们外周系统存在其他组织在运动增强学习记忆的过程中发挥了作用。而骨骼肌作为IGF-1的主要来源之一,且是参与运动的最重要的组织,我们有理由推测骨骼肌是运动后血液中IGF-1增加的主要来源[17]。从以上证据我们可以看出IGF-1以“信使”的身份,负责外周与中枢系统的信息传递,在运动增强学习记忆的过程中起到了重要作用。
1.3 VEGF
VEGF是血管内皮细胞特异性的肝素结合生长因子,传统观点认为,其在机体中的主要作用是诱导血管新生。但近些年的研究发现,VEGF可以使体外培养的大脑皮层脑细胞增殖20%~30%,脑室内注射VEGF也可以引起脑室下区和海马齿状回颗粒细胞神经发生增加,而VEGF基因敲除小鼠的海马齿状回及前脑室下层的神经发生有明显地降低,这都说明VEGF对大脑神经发生也有着重要作用[18,19]。鉴于其对中枢系统结构及功能的作用,研究者对VEGF在运动增强学习记忆中的作用做了探讨,发现运动可以诱导骨骼肌、大脑和肺三类组织中VEGF表达量的增加[20],且VEGF在运动引起的海马区神经发生增加及动物空间学习记忆能力增强中起到了重要作用[21,22]。于是,研究者们推测VEGF可能通过诱发神经发生介导运动对学习记忆的影响[23]。但之后却有研究发现,神经发生在VEGF参与的对学习记忆的增强中并非必要条件[24]。研究者通过在体电生理技术对VEGF在海马区神经可塑性调节的研究中,发现VEGF可以诱发长时程增强(long-term potentiation,LTP),且对这种效应的维持有重要作用,这说明VEGF可能参与了神经元之间突触可塑性的调节[25],甚至可能通过与神经元上受体结合直接参与调节突触的形成[26],具体以何种方式参与突触的调节还有待探究。但以上证据足以证明VEGF在运动增强学习记忆的过程中起到了重要作用,且很有可能是连接外周和中枢的又一重要因子。
2 运动增强学习记忆的分子机制
基于上述运动增强学习记忆相关的三种调节因子研究现状的总结与分析,推测其以突触为结构基础的分子机制如图1。运动增加机体中IGF-1的表达量,其穿过血脑屏障,与神经元和胶质细胞上受体结合激活MAPK和PI3K信号通路,诱导离子通道、神经递质和BDNF等基因的表达,增加突出可塑性,并易化突触形成,增强学习记忆[27,28]。VEGF可通过与IGF-1相似的方式参与运动对学习记忆能力的增强,运动可引起骨骼肌等组织VEGF分泌量增加,其随着循环系统,穿过血脑屏障到达中枢。VEGF除通过促进中枢系统血管再生,增加中枢系统执行功能外,还可与胶质细胞或内皮细胞上Flk-1受体结合,诱导BDNF等相关因子表达量的增加,进而增加突触可塑性。另外,VEGF还可能与神经元上受体结合,直接参与调节突触的形成及可塑性的变化,从而增强学习记忆。此外,运动引起BDNF的基因表达增高后,首先以ProBDNF的形式分泌至细胞外,并由tPA完成剪切产生BDNF,BDNF与突触前膜的TrkB受体结合,提高突触素和突触蛋白-I的合成,促进突触前膜神经递质的分泌,增加突触后膜Ca2+内流,易化晚期LTP形成[29]。而BDNF与突触后膜TrkB受体结合不仅可以激活AMPAR和NMDAR,易化LTP的形成,同时可以通过激活PI3K及MAPK信号通路,促进下游离子通道、神经递质和生长因子等基因的表达,进而促进晚期LTP的形成,提高突触可塑性,增强学习记忆。另外,近些年研究发现,TrkB受体的增加或与长期记忆增强有关[30],这提示我们进一步全面揭示运动增强学习记忆的研究,除了关注BDNF、IGF-1及VEGF等调节因子本身外,其相关受体也不容忽视。
3 动物与人类相关研究的比较分析
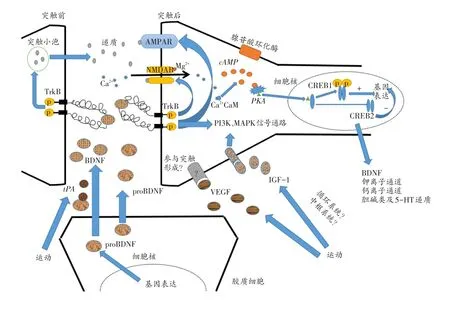
图1 运动增强学习记忆以突触为结构基础的可能的分子机制
随着运动增强学习记忆的动物研究不断深入、人类脑成像等技术的不断发展,运动对人类中枢系统增益的研究在借鉴动物相关研究结果的基础上也逐渐发展起来,成为该领域研究的热点[31-34]。虽然,研究表明运动对人类和动物的认知功能有相似或相同的积极作用,但因为人类和动物的差异性,两类研究在方法及结果上存在许多不同之处[35-37]。首先,动物和人类运动模型不同,主要表现在运动的方式和周期上。动物运动的方式为自主运动、强迫运动和非自主运动,主要由转轮、跑步机和电刺激仪来完成,运动周期一般在1~2个月。而人类的研究多为骑车、散步、耐力训练和协调性训练等体育训练的方式,运动方式多样化且多为温和舒适的运动,周期1个月至2年,周期较长。此外运动量的控制上,动物研究一般不会根据最大摄氧量单独为实验动物设计实验方案,但人类研究会根据自身条件为被试提供尽可能合适的运动量。但无论动物还是人类研究,结果都表明不同的运动方式及运动量对中枢系统的影响有所不同[36,38-41],这就要求研究者在今后的研究中对不同运动模型做更为细致的研究和探讨。其次,动物研究和人类研究关注的认知类型及测试方法不同,动物研究多关注海马相关空间记忆及情绪记忆等不同记忆类型,测试方法为水迷宫、Y迷宫、条件性恐惧记忆、被动回避及新物体识别测验等,相对简单。而人类相关研究除了知觉测验、智商测试、词表学习测验、图像记忆测验、空间记忆测验等心理学学习记忆测试外,还涉及前额叶相关的执行功能、注意控制、逻辑推理能力等高级认知功能,并且有些研究者也试图直接将学业绩效、学业发展水平、工作表现等现实生活中的表现与运动联系起来,测试方法多为心理学相关研究范式,研究跨越认知神经科学、基础心理学、社会心理学、计算机科学等多领域、多层次[34,37]。另外,因为人类研究本身的限制,只能采用脑成像技术(ERP、fMRI、MRS、TMS)等非侵入式手段对运动影响学习记忆做较为宏观层面的研究,而动物研究则可以对运动增强学习记忆的内在机制做细致的研究。总之,动物研究和人类研究都以自己的方式不断开展,在揭示运动增强学习记忆的内在机制及相关应用研究中都具有重要意义。
4 总结与展望
综上所述,一方面,随着运动增强学习记忆的动物研究不断深入,运动增强学习记忆的以突触为结构基础的分子机制已较为详尽。但现今研究更多地关注了运动后中枢系统的变化,对运动直接引起的机体外周系统的变化却少有问津,致使目前对于运动增强学习记忆的上游机制知之甚少。另外,胶质细胞作为外周系统与中枢系统神经元联系的重要枢纽,在上述分子机制中也起到相当重要的作用,且越来越多的研究证明胶质细胞可以直接参与调节神经元之间的活动[42-45]。因此,运动增强学习记忆的进一步研究,需要给予运动直接影响的外周系统和胶质细胞更多关注。这有助于我们更加全面地认识运动对中枢系统的作用,了解运动的实效性。另一方面,随着脑成像等技术的发展,人类运动增益的相关研究也与动物研究同步进行,虽然两者之间在研究方案和方法上存在许多差异,但却得到了较为一致的结果,两者相辅相成,帮助我们更加全面地认识运动给个体带来的积极作用。随着相关技术的不断发展与成熟,可以将动物研究与人类研究相结合,使得动物研究结果能更好地指导人类研究的开展,为人类相关应用类研究提供更多的支持和帮助,从而充分发挥该领域研究的现实意义。
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07 06:21:30
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
自然杂志(2021年6期)2021-12-23 08:24:46
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
汽车维修与保养(2021年8期)2021-02-16 00:28:20
现代装饰(2018年5期)2018-05-26 09:09:01
工业设计(2016年4期)2016-05-04 04:00:15
西南医科大学学报(2015年1期)2015-08-22 13:01:48
电源技术(2015年5期)2015-08-22 11:18:38
