
山东山旺中新世硅藻页岩中植物化石研究新材料
2018-06-22 01:12山东科技大学山东省沉积成矿作用与沉积矿产重点实验室山东青岛66590山东科技大学地球科学与工程学院山东青岛66590
山东科技大学学报(自然科学版) 2018年4期
,, ,,(.山东科技大学 山东省沉积成矿作用与沉积矿产重点实验室,山东 青岛 66590; .山东科技大学 地球科学与工程学院,山东 青岛 66590)
山旺盆地位于山东省临朐县城东北22 km (118°21′E, 36 54′N) 处(图1),在大地构造位置上,该盆地位于欧亚板块东缘郯庐断裂带上,新近系自下而上为牛山组、山旺组和尧山组。山旺组以硅藻页岩为主,是中国北方新近系中新统的典型剖面[1-2]。山旺古生物化石丰富,保存精美,在硅藻页岩岩层中埋藏着难以数计的花、鸟、鱼、虫、兽等精美动植物群[3]。山旺植物群是我国新生代植物群中研究程度最高的代表,主要以温带落叶植物为主,并混生了部分亚热带常绿和落叶阔叶植物,1978年出版的《中国新生代植物》就记录和报道了总计43科87属125种[4]。截止2001年,山旺植物群可靠记录 44 科 88 属128 种,其中苔藓植物1科1属1种、蕨类植物1 科1 属 1未定种、裸子植物 1 科 2 属 2种(含 1未定种)、被子植物 41 科 84属 124 种[5-7]。八十年代以前,山旺植物群的研究工作主要侧重于外部宏观特征的描述,对内部细胞组织结构的解剖学特征研究不足,制约了类群演化生物学的深入研究和对该植物群性质的理解。王宇飞[8]运用国际古植物研究的新方法、新技术,如角质层分析、叶结构分析等,将植物外部宏观特征与微观结构相结合,开展对山旺重要植物类群如杜仲属、榉属、钱耐属和油杉属球果化石等的研究,取得了重要进展,建立了化石新属钱耐属[5],成功开展了古环境定量重建的研究。本次研究发现了保存完好的植物叶化石和球果化石,新发现的珍贵材料对于认识山旺植物群的区系特征与性质可以提供可靠证据。借鉴前人先进的方法,将植物体外部形态学与内部细胞结构研究相结合,并与现生最近亲缘植物进行对比分析,提高了化石植物鉴定的可靠性,提供了现代植物微观细胞结构的实验数据,认为植物化石为中华栎(QuercussinomiocenicaHu et Chaney,1940),确认山旺植物群中松属球果的存在,为研究山旺植物群、古气候和中新世松属的分布与演化提供了重要新材料。
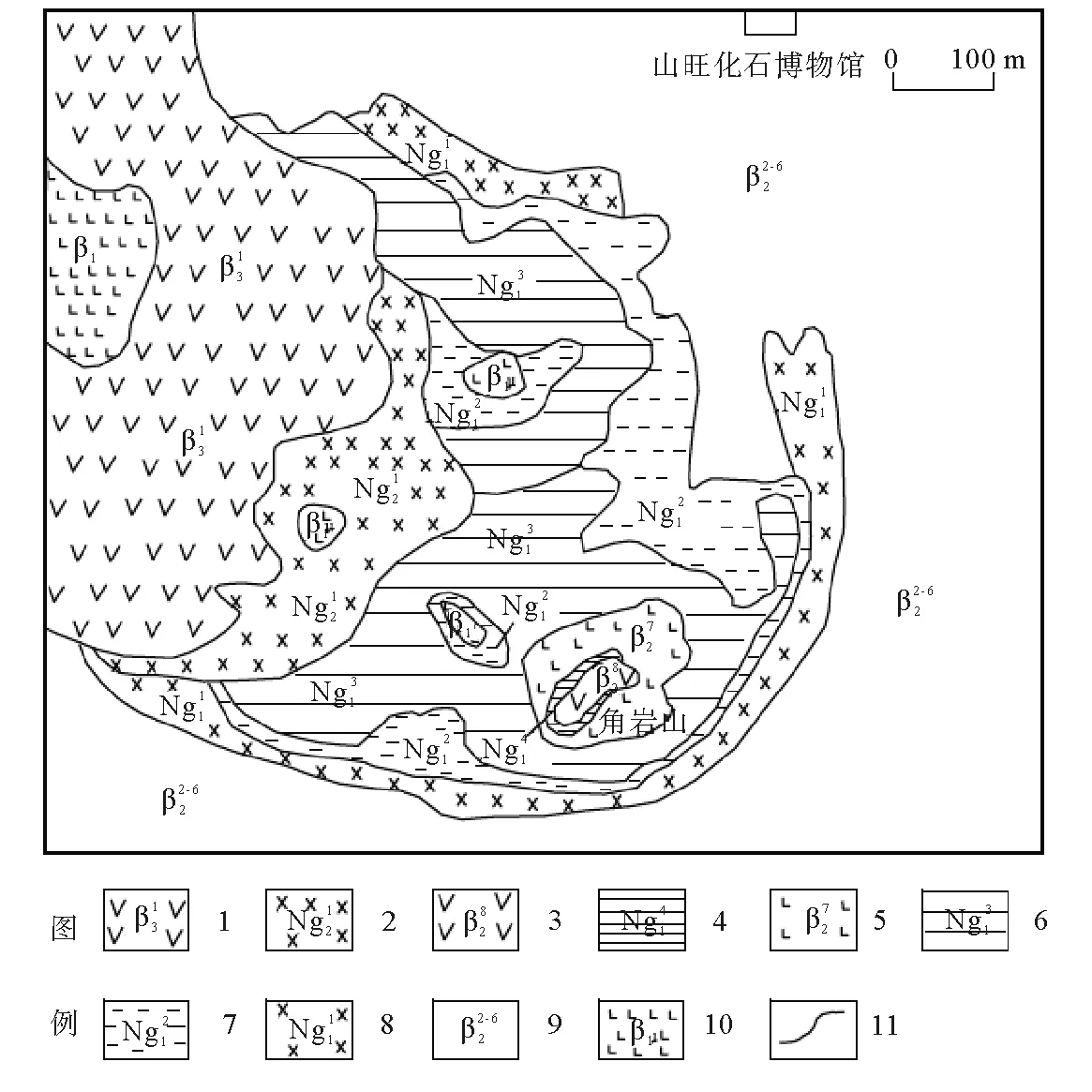
图例说明:1-2—尧山组:1—碱性橄榄玄武岩,2—火山碎屑岩;3-8—山旺组:3—碱性橄榄玄武岩,4—黑色页岩,5—碧玄岩,6—黄绿色泥岩,7—硅藻质页岩,8—火山碎屑岩;9—牛山组玄武岩;10—浅成-超浅成侵入岩;11—地质界线图1 山旺盆地地质略图[9]Fig.1 The geological map of Shanwang basin
1 材料与方法
1.1 实验材料
本文植物化石均采集于山旺组硅藻页岩的顶部,距离剖面顶部约1.3 m的深灰色泥岩层中(图2),野外采样编号第5层。为了研究植物叶表皮结构,并与现生植物进行对比,于中国科学院植物研究所北京植物园采集了现生植物麻栎(Quercusacutissima)、锐齿栎(Quercusacutidentata)和板栗(Castaneamollissima)叶片,并分别进行了叶表皮的分析。

图2 山东省临朐山旺组硅藻页岩地层柱状图Fig.2 Histogram of diatomaceous shale of Shanwang formation at Linqu, Shandong province
1.2 试验方法
1.2.1 植物叶片角质层的化学处理
植物角质层的处理方法多种多样[10-11],本文对植物化石和现代植物叶化石的处理方法如下:
化石植物叶片处理方法:
1) 在植物化石中部靠近叶脉处取一小块放入蒸馏水中浸泡12 h;
2) 放入25%的HCl中浸泡 9 h,之后放入蒸馏水中反复洗涤至中性;
3) 转入塑料瓶中,加入36%HF浸泡约 12 h以上,待岩石完全溶解,获得角质层;
4) 将角质层用蒸馏水反复洗涤至中性,然后转入称量瓶并加入浓度65%的HNO3,待角质层颜色褪为浅棕色后,用吸管吸出 HNO3,用蒸馏水洗涤至中性,并在蒸馏水中浸泡 0.5~1 h;
5) 加入5%的 NH3·H2O 1~2 滴浸泡至半透明后,将上下表皮分开,再用蒸馏水洗涤成中性;
6) 将角质层移入培养皿中,展平于载玻片上,盖上盖片,置于镜下观察、拍照。
现生植物叶片处理方法:
1) 将植物叶片中部刮去表皮毛,放入蒸馏水中清洗;
2) 用剪刀在叶片上取材,在叶片中部靠近叶脉处取出一部分,用蒸馏水清洗;
3) 将取出的叶片放在试管或烧杯中,按1∶1比例加入30%浓度的双氧水与冰乙酸,在70~80 ℃水浴中加热,待叶片颜色呈白色,上下表皮分开即可;
4) 取出样品,用蒸馏水清洗至中性,放在培养皿中用解剖针剥离角质层,平展于载玻片上,盖上盖片,置于镜下观察、拍照。
2 标本描述
2.1 植物叶标本描述
2.1.1 化石标本描述
本次发现植物化石有单个叶化石,还有小枝化石,枝化石上有20余叶片,叶披针形或线状披针形,长9~15 cm,宽1.5~3.7 cm(图3)。顶端长渐尖,基部圆楔形,两侧对称。叶柄粗短,长约0.5 cm。叶缘具疏齿,齿尖具刺芒,长约1 mm。中脉强壮;侧脉11~18对,与中脉夹角45°~60°,位于叶基部的夹角更大,达缘脉序;三次脉较密的连接于侧脉间,细脉明显成网状。与现生壳斗科栎属麻栎叶形最为相似(图3-1,2,4)。
气孔器描述:气孔器长约30 μm,宽约20~23 μm,气孔器排列密集,略凸起于表面,排列无规则,随机分布(图4-1,2)。
2.1.2 化石与现生植物叶标本对比
栎属植物叶片全缘或锯齿,叶形变化较大,其中全缘叶的脉序为羽状弧曲脉,而锯齿的叶片脉序多为羽状达缘脉,侧脉排列较为整齐,三次脉常与侧脉垂直[4]。本文研究的植物叶经与现生壳斗科栎属的麻栎、锐齿栎以及栗属板栗植物比较(表1),发现化石标本叶结构特征以及表皮结构特征(图4),与栎属麻栎植物更为相似,与栗属植物差别较大。再将化石标本与胡先彇和钱耐[12]命名的化石标本中华栎(QuercussinomiocenicaHu et Chaney,1940)相比较,发现该化石标本与中华栎特征极为相符,所以将其鉴定为中华栎(QuercussinomiocenicaHu et Chaney,1940)。现生植物叶形态描述如下:
麻栎QuercusacutissimaCarruth
叶外表结构描述:叶片形态多样,通常为长椭圆状披针形,长8~19 cm,宽2~6 cm,顶端长渐尖,基部圆形或宽楔形,叶缘有刺芒状锯齿,叶片两面同色,幼时被柔毛,老时无毛或叶背面脉上有柔毛,侧脉每边13~18条;叶柄长1~5 cm[13]。
表皮结构描述:上表皮无气孔器;表皮细胞为多边形,四边形至六边形不等,垂周壁平直,细胞直径多在12~23 μm;脉络细胞与一般表皮细胞有差异,多为规则长方形,彼此平行整齐排列,长短不一,长度多在10~36 μm,宽度多在5~11 μm;由多列脉络细胞组成的主脉络清晰,次脉络亦可见;叶脉上偶见多细胞毛基。下表皮具无规则型气孔器,长约20~29 μm,宽约16~26 μm,气孔器排列密集,略凸起于表面,排列无定向, 随机分布在脉络构成的网格内;表皮细胞为不规则多边形,四边形至七边形不等,有的近圆形,细胞垂周壁较平直,直径多在8~20 μm,整体略小于上表皮细胞;脉络细胞多为较规则长方形,彼此较平行整齐排列,长短不一,长度多在14~40 μm,宽度多在6~12 μm,彼此平行排列构成叶脉,连接成网格状。
锐齿栎Quercusalienavar.acuteserrataMaxim
叶外表结构描述:叶长椭圆状倒卵形至倒卵形,长10~20(~30) cm,宽5~14(~16) cm,顶端短渐尖,基部楔形或圆形,叶缘具粗大锯齿,齿端尖锐,内弯,叶背密被灰色细绒毛,侧脉10~15对,有时更多,叶柄长1~3 cm,无毛[13]。
表皮结构描述:上表皮无气孔器;表皮细胞为多边形,四边形至六边形不等,垂周壁平直,少数浅波状,细胞直径多在15~28 μm;脉络细胞多为较规则矩形,彼此平行整齐排列,长度多在17~30 μm,宽度多在11~16 μm;由多列脉络细胞组成的主脉络清晰,次脉络亦可见,叶脉彼此连接成网格状;具单细胞表皮毛,毛基稀疏随机分布。下表皮具无规则型气孔器,长约16~22 μm,宽约13~17 μm,气孔器排列密集,略凸起于表面,排列无定向,随机分布在由脉络构成的网格内;具单细胞表皮毛,表皮毛长而分布密集。
板栗CastaneamollissimaBl.
叶外表结构描述:叶革质,椭圆至长圆形,长11~17 cm,宽度达7 cm,顶部短至渐尖,基部近截平或圆,或两侧稍向内弯而呈耳垂状,常一侧偏斜而不对称,新生叶的基部常狭楔尖且两侧对称,叶背被星芒状伏贴绒毛或因毛脱落变为几无毛;叶柄长1~2 cm[13]。
叶表皮结构描述:上表皮无气孔器;表皮细胞为多边形,四边形至七边形不等,垂周壁平直,细胞直径多在3~8 μm;脉络细胞多为较规则长方形,彼此平行整齐排列,长度多在5~11 μm,宽度多在3 μm左右;由多列脉络细胞组成的主、次脉络清晰,彼此连接成网格状;具多细胞毛基,毛基部的细胞一般特化成辐射状,毛基直径多在6~10 μm,稀疏的随机分布。下表皮具圆形或近圆形无规则型气孔器,长约16~19 μm,宽约15~17 μm,气孔器排列密集,略凸起于表面,排列无定向,随机分布在由脉络构成的网格内;表皮细胞为不规则形,四边形、五边形、六边形和七边形等,细胞垂周壁较平直,少数略有起伏,直径多在6~13 μm,略大于上表皮细胞;脉络细胞多为规则长方形,彼此平行整齐排列,长短不一,长度多30~50 μm,宽度多8~11 μm,多列脉络细胞彼此平行排列构成叶脉,连接成网格状。

表1 研究标本与现生植物及中华栎化石特征比较Tab.1 Comparisons of characteristics between fossil specimens, living plants and Quercus sinomiocenica Hu et Chaney
2.2 球果标本描述
松属球果一般由若干个螺旋状紧密排列的种鳞复合体构成,并且种鳞先端膨大,具有鳞盾和鳞脐[14],松属球果种鳞结构如图5。
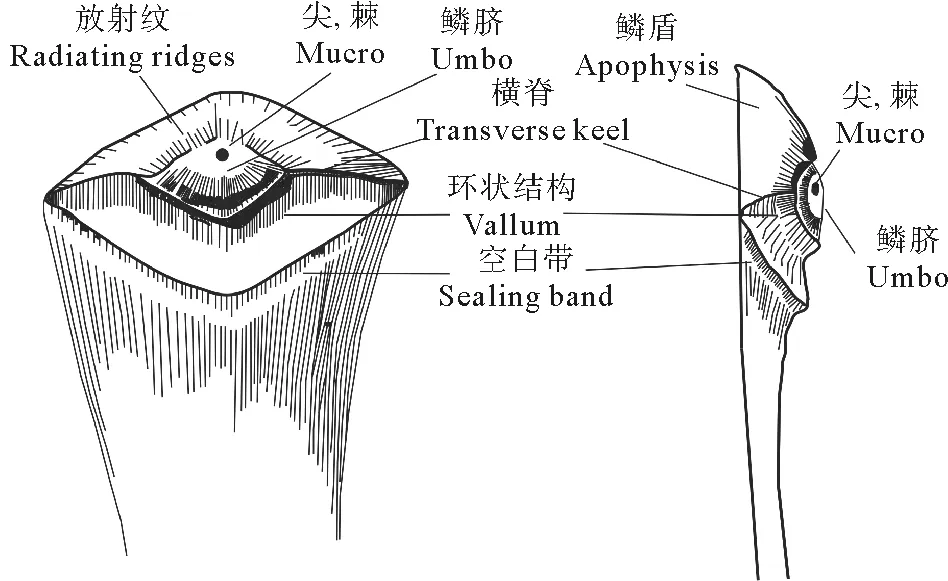
图5 松属球果种鳞结构素描图(修改自Xing et al. 2010[15])Fig.5 Sketch of seminiferous scale structure of Pinus cone
2.2.1 化石标本描述
球果化石个体较小,卵圆形,长16 mm,最宽处约12 mm,由种鳞螺旋状排列组成;种鳞先端膨大,鳞盾菱形,鳞脐背生微凹,椭圆形,位于鳞盾中央,具不明显的横脊,种鳞未开裂(图6)。
2.2.2 现生标本与化石标本对比
黑松Pinusthunbergii
球果圆锥状卵圆形或卵圆形(图6),长4~6 cm,直径3~4 cm,有短梗,向下弯垂;中部种鳞卵状椭圆形,鳞盾微肥厚,横脊显著,鳞脐微凹,有短刺[14]。
本文球果化石与化石和现生松属植物的球果进行比较[14-16],发现其与黑松和马尾松的球果最为相似。与未成熟开裂的黑松球果大小相似,但由于球果化石保存不是特别完整,故无法进一步鉴定到种。所以最终判断该球果化石为松科松属(PinusLinn.1753)。
3 地理分布演化及古气候
壳斗科是北半球热带、亚热带和温带森林的主要树种,是最大的、最重要的木本植物之一,共8属1047种[17]。我国的壳斗科植物约300余种(含特有种164种),占世界总数的三分之一以上,其中栎属(包括青冈属)104种[18]。栎属是北半球亚热带常绿阔叶林、硬叶常绿阔叶、高山灌丛及温带落叶阔叶林的优势成分,在地史上栎属也是分布最广的化石属之一,自始新世就是北半球各植物群的主要和常见分子[19]。通过对栎属化石的详细研究,发现栎属的系统演化规律,解释其区系的发生和发展,对于揭示北半球热带及其温带植物区系的发生和发展及其演化规律具有重要意义[20]。
松属是松科中最大的属,现生植物约110种,主要分布于北半球[21]。国外最早可靠的松属化石记录是早白垩世[22],近来中国科学院南京地质古生物研究所张璐瑾先生[23]通过孢粉分析,在四川威远地区晚三叠世的地层中发现威远松(Pinusweiyuanensis)及环抱松(P.amplexus)2个孢粉化石新种,说明在晚三叠世松属就已经出现在康滇古陆。山旺硅藻页岩中曾经发现过裸子植物松科花粉如油杉属、雪松属、松属、云杉属、落叶松属、铁杉属等[24],本次松属球果化石的发现进一步证实了中新世时期松属植物在山旺地区的存在,因球果化石较大,被风或者流水做远距离搬运的可能性较小,来源应为原地或附近地区。WANG等[25]在山旺发现了油杉属新种Keteleeriashanwangensis,为介于现存的油杉和铁坚油杉之间的中间类型,说明山旺地区松科植物的多样性,反映山旺地区为亚热带山地气候类型。古近纪至新近纪在中国发现松科球果的化石点除山旺之外,还有与山旺基本同期的云南小龙潭组发现的P.prekesiya[15],浙江宁海、天台等地晚中新世嵊县组发现的P.premassoniana、P.specioca、P.preyunnanensissp.nov.[26-27],云南洱源上新世三营组发现的P.yunnanensis[28],河北涞源县斗军湾晚始新世至早渐新世含煤岩系中发现的P.prototabulaeformis[29]。古油松的发现暗示了东亚类群的祖先可能自晚始新世开始出现。这些松属球果化石的发现为研究植物演化与分布提供了宝贵的材料,说明中国中新世时松科植物就具有多样性,分布较为广泛。
化石植物表皮的气孔参数与大气CO2浓度有着相关性,但由于本次做出的气孔数量较少,表皮细胞不够清晰,还需要进一步完善角质层分析方法,达到定量获取古气候参数的要求。前人[30-35]利用古植物及孢粉学对山旺古气候的研究不断丰富,并从定性向定量发展,如叶相分析法、共存因子分析法、孢粉植物群共存分析法、特有种气候分析法、植物分布区叠加分析法等,获得大量古气候参数(表2)。王宇飞等[8,34]利用特有种分析法,根据现有植物杜仲萌发条件,得出山旺古春季的温度为13~20 ℃,高于现在的12.9 ℃。不同方法对于同一化石产地古气候定量分析得出的结论有明显区别,究竟哪种方法更准确,将会随着化石资料的不断丰富,有待于今后更进一步的研究。
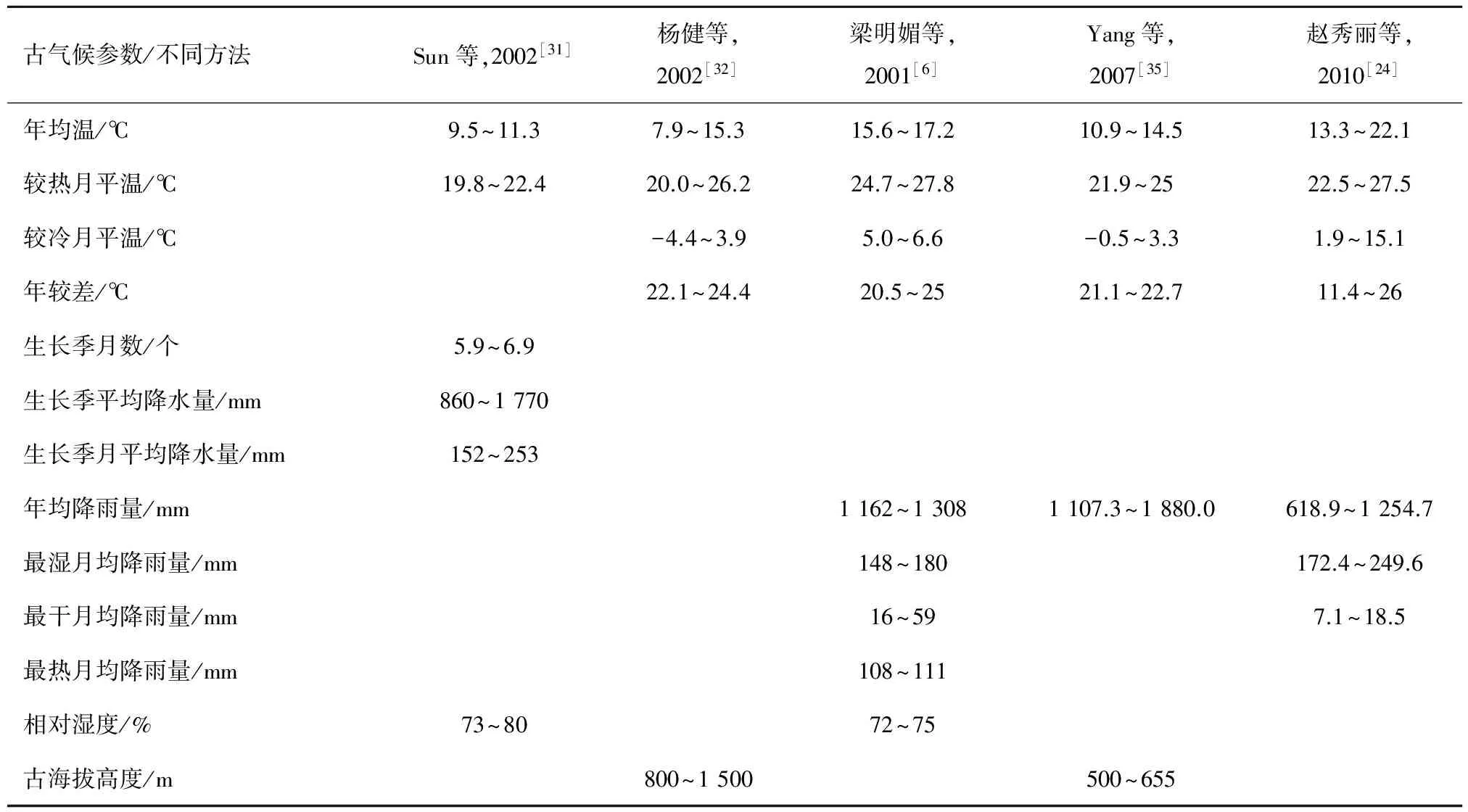
表2 山旺古气候参数对比表Tab.2 The contrast table of palaeoclimate parameters in Shanwang
4 结论
1) 本文对山旺硅藻页岩中新发现的植物叶化石进行研究,与现生壳斗科植物麻栎、锐齿栎和板栗进行叶结构和表皮结构特征的比较分析,将叶化石鉴定为中华栎(QuercussinomiocenicaHu et Chaney,1940)。
2) 叶结构特征和表皮结构特征相结合可以作为壳斗科各属之间的划分依据,通过化石与现生栎属植物表皮结构的对比,提高了鉴定的准确性,提供了现生植物细胞结构的实验数据。
3) 将球果化石与现生松科各属球果进行比较,鉴定为松属,进一步证实了山旺地区中中新世松属的存在,为深入研究古气候提供了依据。
4) 两种植物化石的发现证实了山旺地区中中新世阔叶植物与针叶植物共存的面貌,反映了该地区亚热带山地气候类型。
参考文献:
[1]孙博.山旺植物化石[M].济南:山东科学技术出版社,1999:1-167.
[2]覃文圣,刘建波,韩宝福,等.山东山旺中新世硅藻土的纹层类型及成因[J].沉积学报,2004,22(2):267-275.
QIN Wensheng,LIU Jianbo,HAN Baofu,et al.Types and origin of diatomaceous laminae of the miocene Shanwang Formation in Linqu,Shandong province[J].Acta Sedimentologica Sinica,2004,22(2):267-275.
[3]吴增祥.尘封的化石书卷山旺化石[J].走向世界,2005(7):74-79.
WU Zhengxiang.The Shanwang fossils with the passage of time[J].Openings,2005(7):74-79.
[4]中国新生代植物编写组.中国新生代植物[M].北京:科学出版社,1978:3-200.
[5]WANG Y F,MANCHESTER S R.Chaneya,a new genus of winged fruit from the Tertiary of North America and Eastern Asia[J].International Journal of Plant Sciences,2000,161(1):167-178.
[6]梁明媚,王宇飞,李承森.山旺中新世植被演替及古气候定量研究[J].古地理学报,2001,3(3):11-20.
LIANG Mingmei,WANG Yufei,LI Chengsen.Palaeovegetation succession and quantitative palaeoclimate reconstruction of the Miocene Shanwang assemblage[J].Journal of palaeogeography,2001,3(3):11-20.
[7]叶美娜.关于化石角质层的研究和技术处理方法[C]∥中国古生物学会第十二届学术年会论文集.北京:科学出版社,1981:170-179.
[8]王宇飞.山东山旺中新世植物重要类群演化生物学与古气候定量重建的研究[D].北京:中国科学院植物研究所,2004:47-51.
[9]李泰来,赵弢.山旺古火山口的发现与山旺化石的形成环境[J].现代地质,1988,2(4):91-96.
LI Tailai,ZHAO Tao.Discovery of Shanwang palaeovolcanic orifice and its significance for the preservation of fossils[J].Geoscience,1988,2(4):91-96.
[10]孙柏年,闫德飞,解三平,等.化石植物气孔与碳同位素的分析应用[M].北京:科学出版社,2009:48-61.
[11]孙柏年,石亚军,张成君,等.植物化石角质层分析及其应用[M].北京:科学出版社,2005:8-21.
[12]HU H H,CHANEY R W.A Miocene flora from Shandong province,China[M].Washington:Carngie Institute of Washington Publication,1940,1-147.
[13]中国科学院中国植物志编辑委员会.中国植物志第二十二卷[M].北京:科学出版社,1998:210-270.
[14]中国科学院中国植物志编辑委员会.中国植物志第七卷[M].北京:科学出版社,1998:204-280.
[15]XING Y W,LIU Y S,SU T,et al.Pinusprekesiyasp.nov.from the upper Miocene of Yunnan,southwestern China and its biogeographical implications[J].Review of Palaeobotany and Palynology,2010,160(1-2):1-9.
[16]MILER J C N.Early evolution in the Pinaceae[J].Review of Palaeobotany and Palynilogy,1976,21(1):101-117.
[17]GOVAERTS R,FRODIN D G,KEW R B G.World checklist and bibliography of Fagales (Betulaceae,Corylaceae,Fagaceae and Ticodendraceae)[M].London:The Royal Botanic Gardens,Kew Publishing,1998:201-204.
[18]HUANG C J,ZHANG Y T,BARTHOLOMEW B.Flora of China Vol 4[M].Beijing:Science Press,1999.
[19]周浙昆.栎属的历史植物地理学研究[J].云南植物研究,1993,15(1):21-33.
ZHOU Zhekun.The fossil history of quercus[J].Acta Botanlca Yunnanica,1993,15(1):21-33.
[20]ZHOU Z K.Origin,phylogeny and dispersal ofQuercusfrom China[J].Acta Botanlca Yunnanica,1992,14(3):227-236.
[21]FARJON A.Pines:Drawings and descriptions of the genusPinus[M].2nd ed.The Nether Lands:Koninklijkoe Brilli N V,Leiden,2005.
[22]ALVIN KL.Further conifers of the Pinaceae from the Wealdon Formation of Belgium [J].Mémoires de I′Institut Royal des Sciences Naturelles de Belgique,1960,146:1-39.
[23]张璐瑾.川中晚三叠世孢粉[J].中国古生物志新甲种,1984(8).
[24]赵秀丽,王明镇,宋香锁.临朐中新世山旺组孢粉植物群和古气候研究[J].山东科技大学学报(自然科学版),2010,29(3):1-8.
ZHAO Xiuli,WANG Mingzhen,SONG Xiangsuo.Palynoflora and Paleoclimate of Miocene Shanwang Formation in Linqu[J].Journal of Shandong University of Science and Technology(Natural Science),2010,29(3):1-8.
[25]WANG Y F,XIANG Q P,FERGUSON D K,et al.A new species of Keteleeria (Pinaceae) in the Shanwang Miocene flora of China and its phytogeographic connection with North America[J].Taxon,2006,55(1):165-171.
[26]DING S T,WU J Y,CHEN J L,et al.Needles and seed cones of Pinus premassoniana sp.nov.,and associated pollen cone from the upper Miocene in East China [J].Review of Palaeobotany and Palynology,2013,197(1):78-89.
[27]XU X H,WANG Z X,YANG G L,et al.Twopinusspecies from the upper Miocene in Zhejiang,China and their Palaeobiogeographic significance[J].Review of Palaeobotany and Palynology,2015,68-75.doi:10.1016/j.revpalbo.2015.01.003
[28]陶君容,孔昭宸.云南洱源三营煤系的植物化石群和孢粉组合[J].植物生态学报(英文版),1973,15(1):120-126.
TAO Junrong,KONG Zhaochen.The fossil florule and sporo-pollen assemblage of the Shang-in coal series of Erhyuan Yunnan[J].Acta botanica sinica,1973,15(1):120-126.
[29]陶君容,王庆之.河北涞源县的古油松化石[J].植物分类学报,1983,21(1):108-109.
TAO Junrong,WANG Qingzhi.FossilPinusin Laiyuan Xian,Hebei province[J].Acta Phytotaxonomica Sinica,1983,21(1):108-109.
[30]MOSBRUGGER V,UTESCHER T.The coexistence approach:A method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils[J].Palaeogeography,Palaeoclimatology,Palaeoecology,1997,134(134):61-86.
[31]SUN Q G,COLLINSON M E,LI C S,et al.Quantitative reconstruction of palaeoclimate from the Middle Miocene Shanwang flora,eastern China[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2002,180(4):315-329.
[32]杨健,王宇飞,孙启高,等.中国东部中新世山旺古海拔与古气候的定量研究[J].地学前缘,2002,9(3):183-188.
YANG Jian,WANG Yufei,SUN Qigao,et al.Quantitative studies on palaeoelevation and paleoclimate of Shanwang Miocene basin,east China[J].Earth Science Frontiers,2002,9(3):183-188.
[33]李承森,王宇飞,孙启高.定量分析第三纪以来环境变化的新方法-特有种气候分析法[J].植物学报,2001,43(2):217-220.
LI Chengsen,WANG Yufei,SUN Qigao.Climate analysis of endemic species:A novel method for quantitative analysis of global climate change since Tertiary[J].Acta Botanica Sinica,2001,43(2):217-220.
[34]WANG Y F,LI C S,COLLINSON M E,et al.Eucommia(Eucommiaceae),a potential biothermometer for the reconstruction of palaeoenvironments[J].American Journal of Botany,2003,90(1):1-7.
[35]YANG J,WANG Y F,SPICER R A,et al.Climatic reconstruction at the Miocene Shanwang Basin,China,using leaf margin analysis,CLAMP,coexistence approach,and overlapping distribution analysis[J].American Journal of Botany,2007,94(4):599-608.
猜你喜欢
西北植物学报(2022年10期)2022-11-26
干旱地区农业研究(2022年1期)2022-01-28
乐府新声(2021年1期)2021-05-21
四川师范大学学报(自然科学版)(2021年1期)2021-01-03
当代陕西(2020年21期)2020-12-14
创造(2020年11期)2020-03-19
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
中国林副特产(2019年3期)2019-01-10
现代财经-天津财经大学学报(2016年2期)2016-12-01
