
濒危植物梵净山冷杉的球果与种子性状研究
2022-11-26 04:57:50谭廷鸿吴礼佳许金石王祖华
西北植物学报 2022年10期
谭廷鸿,王 卓,吴礼佳,许金石,王祖华,刘 敏
(1 铜仁学院 农林工程与规划学院,贵州铜仁 554300;2 铜仁学院 贵州省梵净山地区生物多样性保护与利用重点实验室,贵州铜仁 554300;3 西北大学 生命科学学院,陕西西安 710069;4 鲁东大学 生命科学学院,山东烟台 264025)
梵净山冷杉,是世界自然遗产地——梵净山的特有种,为第四纪孑遗植物,被列入《中国植物红皮书》并成为国家一级保护濒危植物,1998 年被IUCN列入《世界自然保护联盟濒危物种红色名录》成为全球重点保护的针叶树种之一[1]。由于梵净山冷杉仅分布于海拔2 100~2 390 m 的山体上部,该区域毗邻山脊,地势陡峭,坡向朝北,夏凉冬冷,霜降频繁,尤其冬季寒潮直侵,加上土层较浅,局部生境脆弱,因而种群不连续成片,多呈小面积碎片化分布[2],且近三十年来不断发现有枯立木或成片死亡植株[3]。由于恶劣的生存条件以及长期孤立的群落演替,导致梵净山冷杉的群体遗传变异性低、成年母树更新较慢、幼树长势较弱,加剧了其濒危态势[3-5]。
截至目前,有关梵净山冷杉的研究主要集中在生境特征、森林类型、种群结构、遗传学特性、土壤成分和濒危机制等方面[1, 4-6]。刘增力等[7]认为梵净山冷杉的分布范围极窄,属于珍稀濒危森林类型,且梵净山冷杉常与南方铁杉和阔叶树组成针阔混交林。生长区局部地段由于阔叶树种的侵入,致使林冠不能郁闭,同时林下苔藓植物及凋落物覆盖较厚,严重阻碍种子接触土壤和萌发[8]。李晓笑等[1]研究表明,梵净山冷杉的种群结构存在波动性,但幼苗不足和幼树死亡率高是种群濒危的重要原因,如果不进行迁地保育,整个种群会因气温持续升高而濒临灭绝。保护遗传学研究表明,梵净山冷杉自交现象严重,不存在种群间基因流动,因而遗传多样性水平较低[9]。此外,张玉武等[10]开展了关于梵净山冷杉的就地育繁和迁地驯化栽培等试验研究,但迄今尚未发现有引种或驯化成功的梵净山冷杉小树甚至幼苗。以上相关研究表明,梵净山冷杉的保育和引种驯化存在较大困难与挑战,关键在于种子到幼苗较低的转化率,导致幼苗和幼树天然更新不足。
研究表明,以有性繁殖为主的植物,其种子发育、成熟以及由种子转化成幼苗的过程是生活史中最关键的环节,直接决定种群能否实现更新和壮大[11-12]。对裸子植物而言,球果和种子是物种繁殖系统的重要特征,它们在强大的选择压力下能表现出较强的适应性[13-14]。同时,球果和种子也是受遗传控制较强的繁殖器官,最具有区分和比较意义[15]。然而,当前尚无关于梵净山冷杉生殖特性尤其是球果及种子性状的研究报道。本研究通过对梵净山冷杉结实母树球果的跟踪观测和定期采集,比较研究不同采种期梵净山冷杉球果和种子的基本性状,揭示其球果和种子的发育和成熟规律,为成熟球果适宜采种期的确定提供参考,同时有助于从繁殖生态学角度解析其濒危机制,为进一步有效开展就地保育和迁地繁育提供理论依据。
1 材料和方法
1.1 球果跟踪观测与采集
梵净山冷杉的球果采自世界自然遗产地——梵净山国家级自然保护区梵净山冷杉原生地(27°46′50″~ 28°1′30″N, 108°35′55″~ 108°48′30″E)的野生种群。在分布区中部最适生的纯林带,选取30 株海拔相近,且树龄为第Ⅳ龄级(20 cm ≤ DBH ≤ 30 cm)的结实母树作为球果跟踪观测和采种对象[2],球果发育的跟踪观测起始时间为2018 年7 月15 日。球果采集日期分别为2018 年8 月25 日、9 月10 日、9 月24 日、10 月10 日和10 月26 日,每期每株母树至少采集3 个球果,每期至少采集90 个球果。
1.2 球果形态性状及重量测定
从同期采集的球果中,随机选取30 个,测定每个球果的长、宽,基径、顶径,鲜重、干重,并计算球果的长宽比、基顶径比和相对含水量。球果的长和宽分别取其最长和最宽部位的测定值[16];球果的基径和顶径分别取第2 轮种鳞对应球果基部和顶部的测量值,测量精度0.1 cm。球果干重以球果自然风干至恒重的数值为测定值,测定精度0.01 g。
1.3 种鳞和种子的性状参数测定
从同期采集的种鳞尚未开放的球果中,随机选取15个,待其自然风干至恒重后,按照球果基部、中部和顶部逐一解剖每个球果,测定单个球果的种鳞层数、种鳞总数量和种鳞总重量;测定单球果的未饱满种子数量和重量、饱满种子数量和重量及种子总数量和总重量,并测定单球果种子饱满率和千粒重[17]。
从球果基部、中部和顶部逐一解剖的种鳞和种子中,分别取各部分的种鳞30 片和纯净的种子30 粒,在解剖显微镜(Olympus, SZX 7)下,测量种鳞长、宽、厚度,种子的长、宽和厚度,以及种翅的长度和宽度。种子、种鳞、种翅的长、宽、厚度分别取最长、最宽和最厚部位的测定值[18],精度为0.01 mm。同时,用万分之一天平分别称量种鳞和种子的重量,测定单片种鳞和单粒种子的重量,测定精度0.001 g。
1.4 数据处理
采用Microsoft Excel 2016 对实验数据进行预处理,并使用GraphPad Prim 5和Canvas X软件进行数据统计分析和制图;采用单因素方差分析(One-way ANOVA)对不同采种期的球果、种鳞和种子的性状参数进行多重比较和差异性显著分析。
2 结果与分析
2.1 梵净山冷杉的球果性状
2.1.1 球果的发育及形态变化通过实地跟踪观测和对比拍摄发现,梵净山冷杉的球果每年7 月中旬开始形成,8 月上旬至10 月上旬为发育关键期,10 月下旬陆续成熟并自然掉落,球果从形成至成熟约3 个月。发育早期即8 月上旬,球果呈圆锥形,颜色深灰色或灰紫色,基部逐渐膨大,雌球花的顶部继续发育,球果长宽比小;发育中期为9 月中旬至下旬,球果卵圆形至近圆柱形,深灰色,雌球花发育完全,基部和顶部宽度继续增加,长宽比也逐渐增加;发育后期为10 月上旬至中旬,球果近圆柱形或圆柱形,灰黑色或棕褐色,长宽比最大(图 1,A~C)。此外,跟踪观测和统计数据显示,梵净山冷杉成年结实母树结果枝的球果结实数量通常1~3 个,且树冠上端尤其是靠近顶端的结果枝结实数量多于树冠下端。

A. 发育早期(8月5日);B. 发育中期(9月10日);C. 发育后期(10月10日)
2.1.2 不同采种期的球果形态特征对不同采种期的球果形态进行了比较分析发现,5个采种期的梵净山冷杉球果外形相似,为卵圆形或近圆柱形,灰黑色或棕褐色;球果表面附着斑块状的白色油脂,油脂面积随采集时间后延而增加(图 2)。统计分析球果的形态参数(图 3)表明,随采种时间后延,10 月 26日采集的球果长、宽、基径和顶径最大,但5个采种期的球果基顶径比(球果基径和顶径之比)差异不显著(P> 0.05);前四个采种期的球果长、宽、基径和顶径增加最明显(P< 0.05),10 月10 日和10 月26 日采集的球果长、宽、基径和顶径差异不显著(P> 0.05);与8 月25 日相比,9 月10 日采集的球果长宽比增加不明显(P> 0.05),9 月10 日、10 月10 日和10 月26 日采集的球果长宽比显著增加(P< 0.05),但后3个采种期间的球果长宽比差异不显著(P> 0.05)。

图2 不同采种期梵净山冷杉球果的形态特征
2.1.3 不同采种期球果的重量及含水量进一步测定分析不同采种期单个球果的鲜重、干重和含水量。数据显示,随采种时间的后延,梵净山冷杉单球果的鲜重和干重显著增加(图 4,A~B),以10 月26 日采集的球果鲜重和干重最大,分别为36.98 g和20.33 g;后四个采种期较8 月25 日球果的鲜重分别增加10.33%、38.18%、52.20%和71.14%,球果的干重分别增加24.44%、79.89%、119.08%和149.24%;从较前一采种期的数据增幅来看,后四个采种期的单球果鲜重增幅依次为10.33%、25.23%、10.15%和12.45%,单球果干重的增幅依次为24.44%、44.56%、21.79%和13.77%。表明梵净山冷杉球果重量的剧增期为发育早期和发育中期,即9 月10 日至9 月24 日,而发育后期和成熟期球果的重量增幅减缓,但成熟期球果重量显著高于发育期。此外,8 月25 日至10 月10 日,球果的相对含水量连续显著降低(图 4,C),然而,10 月10 日与10 月26 日相比,球果的相对含水量差异不显著(P> 0.05)。表明球果的重量和含水量与发育时间呈负相关,球果的相对含水量从发育早期至发育后期持续降低,而发育后期至成熟期趋于稳定。

图中不同小写字母表示不同采种期之间差异显著(P < 0.05)。下同
2.2 梵净山冷杉种鳞的性状特征
2.2.1 不同采种期单球果的种鳞数量和重量测定不同采种期梵净山冷杉单球果的种鳞层数、种鳞数量和种鳞干重(图 5)。结果显示,与8 月25 日相比,后四个采种期的单球果种鳞层数、数量和重量均达到显著差异(P< 0.05),单球果种鳞层数依次增加27.50%、47.50%、68.21%和70.13%,种鳞数量依次增加26.30%、48.91%、59.57%和63.48%,种鳞干重依次增加24.23%、79.61%、107.62%和123.02%。从较前一采种期的数据增幅来看,后四个采种期的单球果种鳞层数的增幅依次为27.50%、15.68%、14.04%和1.14%,种鳞数量的增幅依次为26.30%、17.90%、7.15%和2.45%,种鳞干重的增幅依次为24.23%、44.57%、15.60%和7.42%。可见,单球果种鳞数量和重量在发育早期和发育中期即9 月10 日和9 月24 日增加最明显,在发育后期和成熟期逐渐趋于稳定。
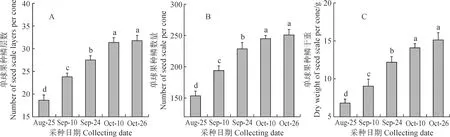
图5 不同采种期梵净山冷杉单球果的种鳞数量和重量
2.2.2 不同采种期球果种鳞的形态特征由于冷杉属球果基部和顶部种鳞不能完全发育,因而对不同采种期球果中部种鳞的形态参数进行了测定,如表1所示。后四个采种期球果中部种鳞的长较8 月25 日均达到显著差异水平(P< 0.05);而对于种鳞的宽,只有10 月26 日采集的球果较8 月25 日达到了显著差异水平(P< 0.05);种鳞的长宽比随种鳞发育逐渐减小,但5个采种期种鳞的长宽比差异不显著(P> 0.05)。对于种鳞的厚度,前3个采种期差异不显著(P> 0.05),直到10 月10 日和10 月26 日才显著增加,较8 月25 日分别增加1.16 倍、1.39 倍。9 月10 日种鳞的干重较8 月25 日差异不显著(P> 0.05),后三个采种期种鳞的干重较8 月25 日显著增加(P< 0.05),分别增加了2.35 倍、3.17 倍和4.76 倍。此外,发育后期和成熟期,即10 月10 日和10 月26 日,球果中部种鳞的厚度和干重差异不显著(P> 0.05)。可见,随着球果的发育和成熟,梵净山冷杉球果种鳞的长、厚度和干重会显著增加,而发育后期和成熟期种鳞的厚度和干重趋于稳定。
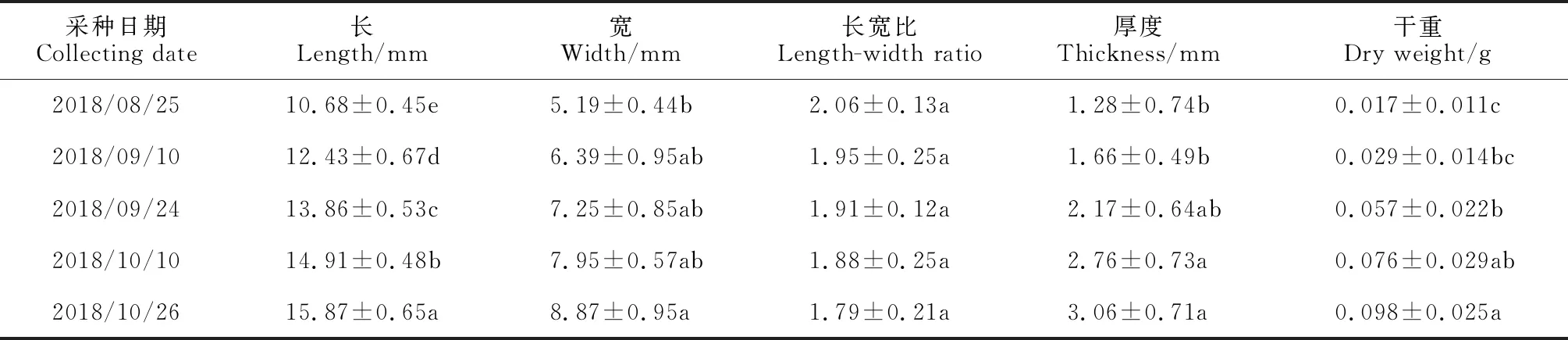
表1 不同采种期梵净山冷杉球果中部的种鳞性状
2.2.3 成熟期球果种鳞的形态特征对成熟期即10 月26 日采集的梵净山冷杉球果的种鳞逐一解剖后,分别选取球果基部、中部和顶部各3 层的种鳞进行拍摄和形态分析。如图 6所示,梵净山冷杉成熟球果的种鳞完全木质化,坚硬。球果基部的种鳞近圆形或方形,中部的种鳞肾形,顶部的种鳞矩扇形。种鳞上下边缘均有细锯齿,下边缘中央凸出形成柄;柄左右两侧不对称。球果基部种鳞的柄较短,中部和顶部种鳞的柄稍长,柄上宽下窄,棕褐色;中部种鳞的柄略宽且厚。种鳞由腹面向背面凹陷,腹面柄两侧分别附着1 粒带种翅的种子,因而种鳞柄两侧的中央区域和下边缘略薄,上边缘略厚;种鳞腹面和背面的中央区域棕褐色,背面的上边缘为黑褐色;腹面粗糙或有颗粒状凸起,背面稍平滑;种鳞腹面顶部中央有刺状突起,较短,质地硬。
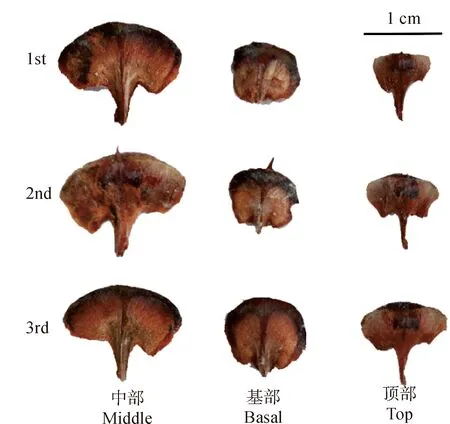
每个部位分别取第1、第2和第3轮的种鳞
2.2.4 成熟期球果种鳞的性状特征进一步测定成熟期球果基部、中部和顶部的第二轮种鳞的性状参数,包括种鳞的长、宽、长宽比、厚度和干重(表 2)。数据显示,球果基部种鳞的长和长宽比最小;顶部种鳞的宽、厚度和干重最小;球果中部种鳞的所有性状指标均为最大。对于种鳞的长,成熟球果基部、中部、顶部之间差异显著(P< 0.05),中部种鳞的长分别是基部和顶部种鳞的2.13 倍、1.85 倍。与种鳞的长类似,成熟球果基部、中部、顶部的种鳞宽差异显著(P< 0.05),中部种鳞的宽分别是基部和顶部的1.21 倍、1.64 倍,基部种鳞的宽是顶部种鳞的1.35 倍。中部和顶部种鳞的长宽比相当,且都显著大于底部(P< 0.05)。基部种鳞的厚度介于中部和顶部之间,但与两个部位的种鳞厚度差异不显著(P> 0.05)。三个部位间的种鳞干重差异显著(P< 0.05),中部种鳞干重分别是基部和顶部的1.52 倍、2.91 倍。
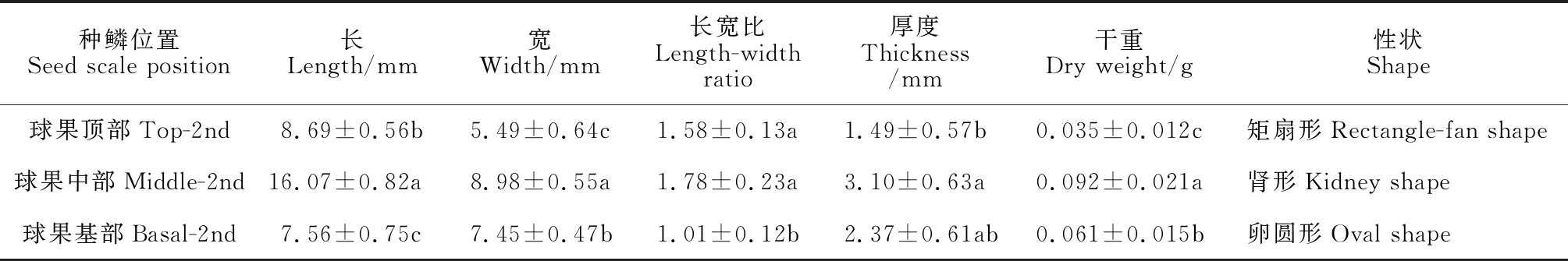
表2 梵净山冷杉成熟球果不同部位的种鳞性状特征
2.3 梵净山冷杉种子的性状特征
2.3.1 不同采种期球果的种子数量对不同采种期梵净山冷杉球果的种子数量进行测定,比较分析单球果未饱满种子数量、饱满种子数量、种子总数量和种子饱满率(图 7)。结果显示,前3个采种期,单球果的未饱满种子数量显著增加,随后两个采种期又显著降低,发育中期即9 月24 日的单球果未饱满种子数量最多。单球果的饱满种子数量、种子总数量和种子饱满率均随采种时间后延逐渐增加,虽然较早两个采种期即8 月25 日和9 月10 日之间无明显差异(P> 0.05),但是后三个采种期较8 月25 日的增长率均显著(P< 0.05),单球果饱满种子数分别增加5.23 倍、13.82 倍、23.57 倍,种子总数分别增加1.87 倍、2.79 倍、3.83 倍,种子饱满率分别增加1.17 倍、2.91 倍、4.19 倍。以上结果表明,发育早期至发育中期,球果以形成新种子为主,而发育中期至成熟期,新种子形成减少,以灌浆增加种子饱满度为主。

图7 不同采种期梵净山冷杉单球果的种子数量及饱满率
2.3.2 不同采种期球果的种子重量对不同采种期梵净山冷杉球果的种子重量进行测定,比较分析单球果的未饱满种子重量、饱满种子重量、种子总重量和种子千粒重(图 8)。数据显示,与单球果的未饱满种子数量类似,单球果的未饱满种子重量在前3个采种期显著增加,随后两个采种期又显著降低,发育中期即9 月24 日最高。单球果饱满种子重量在前两个采种期差异不显著(P> 0.05),后续三个采种期均显著增加(P< 0.05)。单球果种子总重量和种子千粒重随采种时间后延均显著增加(P< 0.05)。与前四个采种期相比,10 月26 日的单球果种子总重量分别增加3.78 倍、2.55 倍、1.31 倍和0.41 倍,种子千粒重分别增加0.87 倍、0.51 倍、0.29 倍和0.13 倍。以上数据表明,梵净山冷杉球果从发育早期至发育中期,种子重量的显著增加主要源于种子数量的增加;而球果发育中期至成熟期,种子的重量增加源于种子数量和种子饱满率的增加。
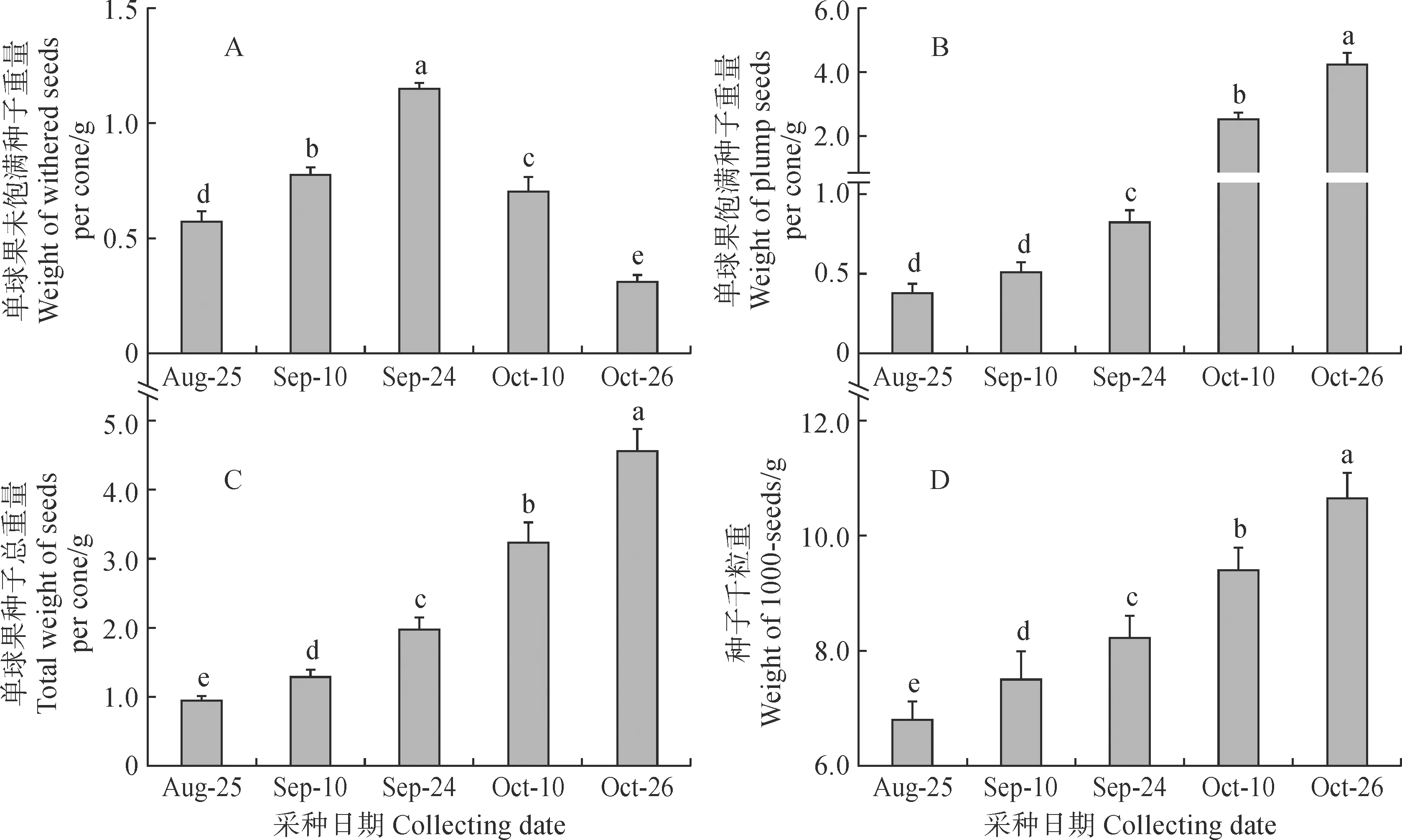
图8 不同采种期梵净山冷杉单球果的种子重量
2.3.3 成熟期球果的种子形态特征对成熟期采集的梵净山冷杉球果进行解剖后,分别选取球果基部、中部和顶部各3 层的种子进行显微拍摄和形态分析。如图 9所示,球果基部、中部和顶部的种子均有明显的种翅,种翅为顶侧生长;种翅上边缘平截,较薄,有锯齿;球果基部和中部的种翅颜色较深、棕褐色,顶部的种翅颜色较浅、棕色;种翅表面有竖条纹和颗粒。种子的种皮与种翅连生,种皮薄,胶质,易碎;种皮表面粗糙,有凹凸纹。种子梭子型,中间宽厚、两端细,表面粗糙,浅棕色;种子含油脂,质地偏软,易撕裂;种子的形态大小依次为球果中部 > 球果基部 > 球果顶部。

每个部位分别取第1、第2和第3轮的种子
2.3.4 成熟球果的种子性状特征进一步测定成熟期球果基部、中部和顶部的种子性状参数,包括种子长、宽、厚度、千粒重,同时测定了种翅的长和宽(表 3)。数据显示,球果中部种子的所有性状参数均为最大,基部和顶部的种子除长度差异显著(P< 0.05)外,其余性状参数无显著差异(P> 0.05)。球果中部种子的长分别是基部和顶部的1.14 倍、1.34 倍,宽分别是基部和顶部的1.28 倍、1.31 倍,厚度分别是基部和顶部的1.39 倍、1.48 倍。球果基部、中部和顶部种子的千粒重差异显著(P< 0.05),中部种子千粒重分别是基部和顶部的1.38 倍、1.60 倍。数据表明,梵净山冷杉成熟球果的种子并非呈上中下均匀分布,球果中部种子的长、宽和饱满度均显著高于基部和顶部的种子。
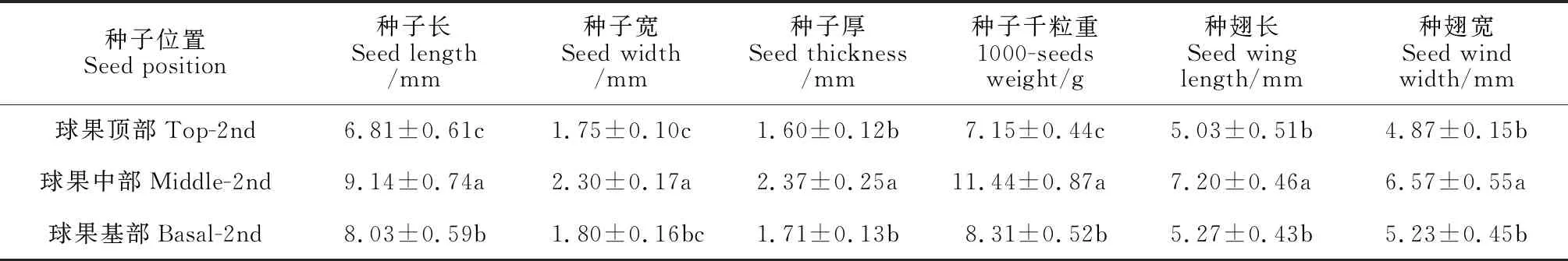
表3 梵净山冷杉成熟球果的种子性状特征
3 讨 论
对于种子植物而言,遗传多样性是生物多样性的决定因素[19],认识遗传多样性和种群结构特征是有效保护濒危植物的重要基础[20-22]。在众多影响物种种群结构的因素中,繁殖系统一直是学者们关注的焦点,因为其表型的多样性程度体现了种群对于环境适应能力的程度[23]。球果和种子是裸子植物的重要繁殖系统,也是冷杉属濒危植物的基因库和种群发展与维持的内在动力[14]。本研究选取梵净山冷杉分布区中部最具物种代表性的适生纯林带的结实母树为采种对象,跟踪观测发现,每年7 月中旬梵净山冷杉的球果开始形成,10 月底成熟并陆续自然掉落。发育早期至发育中期球果的颜色和外形有明显变化,发育后期至成熟期球果的颜色和形态不再发生显著变化。因此,不能以球果的颜色或者形态作为其成熟的判断依据。冷杉属球果的形态发育和成熟过程受到严格的遗传控制[24-25],环境因素对特定物种的种实性状也有影响。影响冷杉属球果遗传分化的生态因子主要是海拔、温度、生长期和光周期,如秦岭冷杉(Abieschensiensis)球果在低海拔发育最好,高海拔处较差[16, 26]。与秦岭冷杉不同,梵净山冷杉分布范围比较狭窄,分布区的坡向基本相同,均为阴坡,不同生境对其种群结构的差异影响不大[1]。但是分布区垂直海拔差约300 m,且不同海拔的小气候和土壤养分不同,因此海拔、土壤和气候等环境因子是否对梵净山冷杉球果发育、成熟及性状特征有显著影响,需进一步研究。本研究发现梵净山结实母树的结果枝能够孕育1 ~ 3 个球果,且单株母树能够满足5 次球果采集,表明当年跟踪的结实母树产果量大于15 个。同属的秦岭冷杉平均结实量为113 个,结实量在海拔高度间、植株间和结实的空间上有差别,而且还存在球果结实大小年现象和生殖间隔期[16-17]。松科植物如油松(PinustabulaeformisCarr.),两年生球果的发育会降低树体结果枝的营养水平,从而引起结实大小年[27],因此我们推测梵净山冷杉球果结实同样存在大小年和生殖间隔。
梵净山冷杉成熟球果的平均长度为7.18 cm,平均宽度3.84 cm,平均鲜重36.98 g,平均干重 20.33 g,单球果平均种鳞层数30.76 层,种鳞数250.67 片。与冷杉属的其他树种相比,梵净山冷杉球果大小居中[16],种鳞数量较多。与球果的长和宽类似,梵净山冷杉单球果种鳞数量和重量在发育早期和发育中期,即9 月10 日和9 月24 日增加最明显,在发育后期和成熟期逐渐趋于稳定,表明梵净山冷杉种鳞与球果的发育和成熟同步,这与松科的大多数树种类似[8, 28]。然而,关于冷杉属种鳞的层数和每层种鳞的数量是否决定球果长和宽,迄今尚无相关报道。梵净山冷杉球果发育中期至成熟期,部分种鳞背面附有油脂,这可能与种鳞密封及种子开放有关[29],此外,梵净山冷杉球果基部、顶部和中部种鳞的形态不同,仅球果中部的种鳞能够完全发育。成熟球果中部种鳞的所有性状指标均最大,每片种鳞腹2 粒种子。与此不同,崖柏球果中部的种鳞通常1 粒种子,其球果基部和顶部的种子不能发育成熟[30]。
冷杉属植物的球果在发育和成熟过程会出现种子败育或发育异常等现象,种子也会存在空粒、涩粒和虫粒[16, 31-33]。本研究发现,随着采种时间的后延,梵净山冷杉单球果的种子数量、种子饱满率和千粒重显著增加;成熟单球果的平均饱满率为82.49%,比冷杉属的其他树种略高;梵净山冷杉成熟单球果的平均种子数为436.67 粒,比同属其他树种的单球果出种量更高[16, 34]。虽然成熟球果基部和顶部的种鳞未能完全发育,却均能正常孕育种子,不过球果上、中、下三个部位的种子形态和性状存在较大差异。有研究表明,在针叶树种中,种子的大小由球果形态所决定[14]。梵净山冷杉球果中部的种子平均长仅9.14 mm,宽2.30 mm,厚2.37 mm,千粒重11.44 g,在同属冷杉树种中偏小[16]。通常,大粒或重量大的种子比小粒种子的贮藏物质更多,种子的大小或重量是植物给每个后代的物质与能量投资指数,大粒种子产出成本高,周期长,种子数量少[35]。梵净山冷杉种子小、重量轻,但是单球果产量高,这是一种长期的适应对策。然而,种子种群是植物一生中有能力移动的关键期,能否保证足够的种子种群经散布后形成土壤种子库,并避开不良环境,在适宜时机达到幼苗的建成,为种群补充后代,对种群的发展和壮大至关重要[36]。虽然梵净山冷杉单球果种子数较多,且饱满率高,但是其分布区域狭窄,生境条件恶劣,林下苔藓和凋落物覆盖较厚,种子成熟后自然散落难以触及土壤,这就极大限制了种子到幼苗的建成。因此,单株对后代的贡献少,难以满足种群的自然更新,且幼树成年周期长,母树结实率衰减;同时,全球气候变暖对针叶树种的繁殖特性造成巨大影响[37],加上冰雹雪凝折枝等自然灾害,使得种群面积越来越小,导致梵净山冷杉逐渐成为濒危树种。
综上所述,不同采种期梵净山冷杉球果、种鳞和种子的性状不同。8 月上旬至10 月上旬为梵净山冷杉球果发育的关键期,期间球果形态和重量显著增加,10 月中旬至10 月底球果、种鳞和种子的形态和性状逐渐趋于稳定,10 月底球果陆续成熟并自然掉落,因此,10 月下旬为成熟球果的最佳采集期。梵净山冷杉成熟球果近圆柱形,灰黑色或棕褐色;种鳞与球果同步发育和成熟,球果上、中、下三个部位的种鳞及种子的性状不同,球果中部的种鳞和种子发育最好。在同属冷杉树种中,梵净山冷杉成熟球果的大小居中,出种量大,种子饱满率高,但是种子小、重量轻,不利于自然散落后形成土壤种子库和实现种子到幼苗的建成。梵净山冷杉球果与种子的上述性状特征,是否成为其致危的决定性因素,仍需进一步研究。
猜你喜欢
山西林业科技(2022年4期)2022-03-12 04:39:48
广西林业科学(2019年2期)2019-07-25 02:28:46
花火A(2019年1期)2019-03-22 02:27:42
中国林副特产(2019年3期)2019-01-10 17:17:11
福建林业(2018年3期)2018-11-07 05:34:36
乡村地理(2018年3期)2018-11-06 06:50:14
铜仁学院学报(2018年7期)2018-09-08 08:50:22
贵茶(2018年6期)2018-05-30 09:53:52
乡村地理(2017年4期)2017-09-18 02:53:52
现代养生·上半月(2017年7期)2017-07-07 21:43:25
