
新型非酒精性脂肪肝模型建立及免疫细胞机制探讨
2018-06-08 09:45陈一平肖百全张景鸿刘位位肖亚强李超乾
中国药理学通报 2018年6期
陈一平,肖百全,张景鸿,刘位位,肖亚强,李超乾
(1. 广西医科大学附属民族医院/广西民族医院老年病科,广西 南宁 530001;2. 广东省生物资源应用研究所药物非临床评价研究中心,广东 广州 510990;3. 广西医科大学第一附属医院急诊科,广西 南宁 530021;4. 广西急诊与医学救援人才小高地,广西 南宁 530021;5. 广西医科大学研究生院,广西 南宁 530021;6. 广西卫生职业技术学院医学系,广西 南宁 530023)
随着肥胖及代谢综合征的流行,非酒精性脂肪肝(non-alcoholic fatty liver disease, NAFLD)的发病率及患病率也呈现逐年增长的趋势[1]。横断面研究显示[2],全球有西方膳食习惯成人中,20%~30%存在肝脏过量脂肪沉积,2%~5%的NAFLD患者存在明显肝脏损害,1%~2%的成人存在患非酒精性脂肪肝炎导致肝硬化的风险。NAFLD明确能使部分肝硬化患者肝功能失代偿,并最终导致死亡或肝移植。另外,NAFLD也造成了国家巨大的经济负担,美国在该病每年花费估计1030亿美元,而英国、德国、法国和意大利花费350亿英镑[3]。我国有研究通过调查6 450名职业人群,发现NAFLD 的患病率为21.71% ,男性为28.68% ,女性为14.47%[4],虽然尚无大数据统计结果,但也能充分说明NAFLD在我国的严峻形势。
为了研究NAFLD的发病机制及治疗方法,制备出接近人类疾病过程,且成功率、重复率高的动物模型显得尤为重要。目前,常用的NAFLD模型建立主要有化学药物诱导、病毒诱导、高脂饲料诱导或转基因及基因敲除动物等方法[5]。众所周知,西方饮食习惯是导致人类肝脏过量脂肪沉积的主要原因[2],膳食因素也是导致人类NAFLD的主要病因[6],所以,高脂饮食诱导是最接近人类发病过程的造模方法。目前,研究对模型动物的选择、高脂饲料的配比及是否应用辅助药物干预等各不相同,但由于小鼠自由摄食存在摄入量不可控,影响模型成功率的缺陷,本研究通过基础饲养联合高脂肠内营养剂灌胃诱导,建立C57BL/6J小鼠NAFLD模型,为该疾病模型的建立及疾病相关研究提供参考和借鉴。
1 材料
1.1实验动物SPF级C57BL/6J♂小鼠,3~4周龄,刚断乳,体质量17~21 g,由湖南斯莱克景达实验动物有限公司提供,实验动物生产许可证号:SCXK(湘)2013-0004。模型制作在广东莱恩医药研究院SPF级动物房观察室完成,许可证号:SYXK(粤)2015-0146。动物房环境温度(20~26) ℃,日温差不超过4 ℃,相对湿度40%~70%,换气次数≥15次/h,12 h照明/12 h黑暗明暗交替,自由摄食摄水。
1.2饲料配方及供能比基础饲料:采用上海普路腾生物科技有限公司小鼠基础饲料,供能比:脂肪12.3%、蛋白21.5%、碳水化合物66.2%。参考李晓冲等[7]高脂饲料配方比例,并加以改进,高脂肠内营养剂配方:20%蔗糖+20%猪油+2%胆固醇+1%胆酸盐。供能比:脂肪69.2%、碳水化合物30.8%。
1.3试剂血清总胆固醇(total cholesterol,TC)批号:16803、甘油三酯(triglyceride,TG)批号:161004、高密度脂蛋白(high-density lipoprotein,HDL)批号:16703、低密度脂蛋白(low-density lipoprotein,LDL)批号:16302、丙氨酸氨基转移酶(alanine aminotransferase,ALT)批号:161206、天冬氨酸氨基转移酶(aspartate transaminase,AST)批号:160904,以上试剂盒均购自浙江伊利康生物技术有限公司;抗小鼠CD4 Percp-cy5.5(批号:550954)、PE标记大鼠抗小鼠IL4(批号:554435)、PE-Cy7标记 小鼠抗小鼠NK-1.1(批号:552878)、Ms CD3e APC 145-2c11(批号:100326),以上流式荧光抗体均购自美国BD公司;胶原酶IV(批号:V900893),购自美国Sigma公司。
1.4仪器FACSVerse型流式细胞仪(美国BD公司);EG1150H型石蜡包埋机+EG1150C型冷台、ST5020型多功能染色机、DM3000生物显微镜(ebp100-04-L荧光)型,均购自德国LEICA公司;病理图像分析系统:DFC450+图像采集软件Leica Application Suite V4,德国LEICA公司;7020型全自动生化分析仪(日本日立公司)。
2 方法
2.1动物分组及模型制备检疫合格的C57BL/6J小鼠经3 d适应性喂养后,按体质量随机均衡地分为2组,即对照组和模型组,每组20只。所有小鼠均自由摄食(基础饲料)及饮水。根据药理实验方法学[8],并结合本实验室既往研究经验,模型组每日上、下午给予高脂肠内营养剂,按每10 g体质量0.3 mL灌胃,对照组给予同样剂量的纯水灌胃。喂养12周及16周分别处死10只小鼠,进行相关指标检测。
2.2肝指数测定用丙泊酚麻醉小鼠后,称其体质量,摘眼球放血后,开腹分离肝脏,观察肝脏大小、颜色、形状,称量肝脏重量,并计算肝指数。肝指数=肝脏重量(g)/体质量(g)。
2.3病理学检测切取适量肝脏组织制作新鲜冰冻切片,再进行油红O染色,2名病理医师在光学显微镜下进行双盲阅片。按照30%以上肝细胞脂肪变性者视为脂肪肝,75%以上肝细胞脂肪变性者视为重度脂肪肝标准得出结论,如存在争议,请第3名病理医师阅片。
2.4血清学指标检测所有小鼠均在早上8点采血,腹主动脉采血分离血清,按照试剂盒操作步骤,应用全自动生化分析仪进行ALT、AST、TC、TG、HDL、LDL等生化指标检测。
2.5流式细胞术检测肝脏NKT细胞及其活性按照本课题组前期研究实验方法[9]稍有改动:用3 mL 40% Percoll重悬小鼠肝细胞的细胞悬液,室温下2 400 r·min-1离心30 min,吸取含红细胞层,加红细胞裂解液3 mL,暗处放置4 min,过滤后,1 500 r·min-1离心5 min,1 mL RPMI 1640细胞培养液重悬,制成肝脏单细胞悬液备用。取肝脏单细胞悬液100 μL,依次加入percp5.5-CD4、FITC-CD69、PECY7-NK1.1、APC-CD3各1.5 μL轻柔混合,暗室室温孵育20 min。PBS液3 mL,1 000 r·min-1离心5 min,弃上清,加入1%多聚甲醛200 μL,于4℃冰箱内固定过夜,24 h内上机检测。CD3+NK1.1+细胞为自然杀伤T细胞(natural killer T cells, NKT cells),CD3+NK1.1+IL-4+细胞为IL-4+NKT细胞,CD3-NK1.1+细胞为自然杀伤细胞(natural killer cells,NK cells),CD3+CD4+细胞为辅助性T细胞(helper T cells,Th cells)。

3 结果
3.1实验动物大体情况各组小鼠造模过程中均未出现死亡,对照组小鼠生长及生活习性正常,模型组小鼠在喂养后期,特别是造模12周以后到16周实验结束时,出现体型稍大、反应迟钝、懒动表现。
3.2小鼠体质量、肝指数变化对照组肝脏呈鲜红色,边缘锐利。模型组12周时肝脏呈现色泽变黄、油腻感,16周上述表现加重,并出现肝脏体积增大,边缘变钝现象。Tab 1结果显示,模型组体质量较对照组有增加,但差异无显著性(P>0.05);肝指数较对照组明显增加,以16周时更加明显(P<0.05)。
3.3血清学指标的变化Tab 2、3结果显示,与对照组相比,模型组ALT及AST水平在造模12周时差异无显著性(P>0.05),16周时出现明显差异(P<0.05);与本课题组之前研究结果类似[10],模型组TC水平在12周明显升高(P<0.05),16周时差距有所减小,但差异仍有显著性(P<0.05);TG水平在12周时降低,但差异无显著性(P>0.05),16周时差异进一步缩小;HDL水平在12周及16周时,降低均不明显(P>0.05);LDL在12周时明显升高(P<0.05),16周时差异更加明显(P<0.01)。
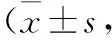
Tab 1 Changes of body weight and liver index n=10)
*P<0.05,**P<0.01vscontrol
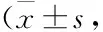
Tab 2 Changes of liver function and blood lipid level in 12 weeks n=10)
*P<0.05,**P<0.01vscontrol
Tab 3 Changes of liver function and blood lipid level in 16 weeks n=10)
*P<0.05,**P<0.01vscontrol
Tab 4 Changes of different lymphocytes n=10)
*P<0.05,**P<0.01vscontrol
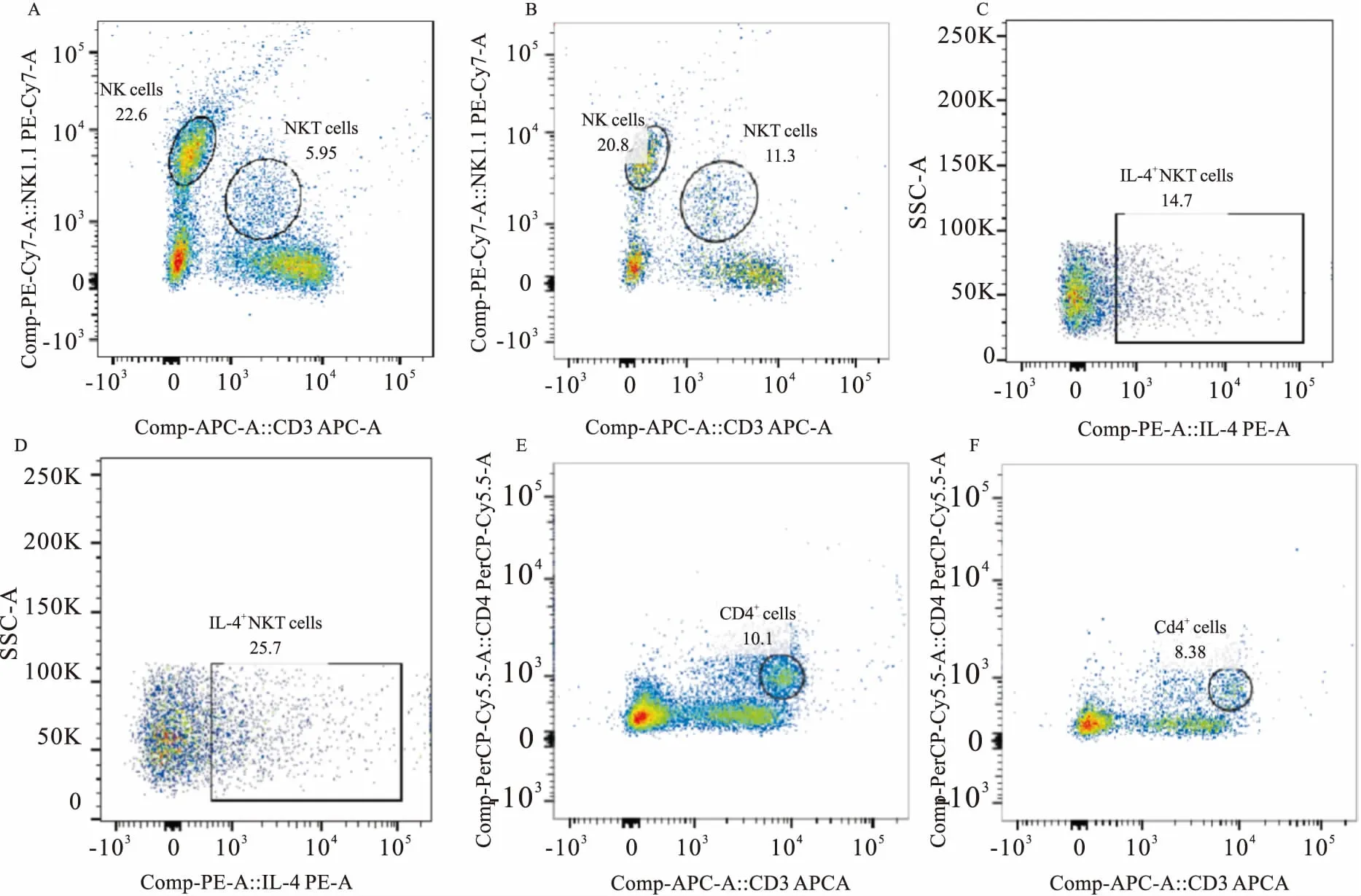
Fig 1 Changes of different lymphocytes
A:Control NK cells and NKT cells; B: Model NK cells and NKT cells; C: Control IL-4+NKT cells; D: Model IL-4+NKT cells; E: Control Th cells; F: Model Th cells.
3.4肝脏不同淋巴细胞变化如Fig 1、Tab 4所示,与对照组相比,模型组NKT细胞比例升高,差异具有显著性(P<0.05),IL4+NKT细胞比例明显升高(P<0.01),而NK细胞比例有升高趋势,Th细胞比例有下降趋势,但差异均无显著性(P>0.05)。
3.5肝脏病理学检查结果如Fig 2所示,16周时,对照组也有少量肝细胞脂肪浸润,模型组脂肪浸润明显,脂肪变性的肝细胞明显超过75%,与造模12周的小鼠肝脏病理相比,肝细胞脂肪变性数量及程度都更加严重。但是,不论是在12周,还是16周,模型组肝脏纤维化表现均不明显。
4 讨论
小鼠NAFLD模型模拟人类因长期高脂高糖膳食,导致过量脂肪在肝脏沉积引起脂肪肝的疾病过程。常见膳食诱导方法是高脂高糖饲料[5]或者蛋氨酸/胆碱缺乏饲料(methionine -choline deficient diet,MCD)[11]喂养小鼠,或联合应用丙基硫氧嘧啶抑制胆固醇分解,增加脂质在肝脏中沉积,或联合应用四氯化碳加重肝脏损害,缩短成模时间等方法[12]。但是单纯高脂高糖饲料喂养,温度、湿度、气压、光照条件等的细微变化,以及高脂饲料本身因为质地较软不利小鼠磨牙等原因,都可以造成小鼠摄食量改变,影响最终模型的成功率。而使用特殊制作饲料或者联合化学药物提高模型成功率的方法,与人类NAFLD的发病过程始终存在较大差异。本研究中,应用高脂高糖肠内营养剂灌胃保证小鼠摄食量,同时以基础饲料自由进食及磨牙棒使用尽可能减少对牙齿的影响。为了避免长期灌胃造成小鼠食道及气道损伤,本研究实验操作人员均为广东莱恩医药研究院长期从事药物毒理研究的熟练实验员,实验过程中,无1只小鼠因灌胃操作出现死亡的现象。
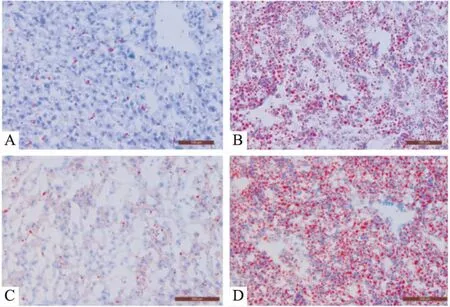
Fig 2 Photomicrographs of liver in mice(frozen section, red oil O×200)
A: Control 12 weeks; B: Model 12 weeks; C: Control 16 weeks; D: Model 16 weeks.
本研究终点,模型组小鼠并未出现体质量明显增加,原因考虑为在模型建造初期,每10 g体质量0.3 mL剂量的高脂肠内营养液导致小鼠出现活动明显减少、精神萎靡、体温下降、摄食减少等反应,而同样剂量饮用水灌胃的对照组小鼠未出现上述现象。虽然后面立即将灌胃量减半(每10 g体质量0.15 mL剂量),后逐步增加至预计灌胃量,小鼠精神、活动、体温、摄食情况逐渐恢复,但由于上述应激反应发生是在小鼠断乳后体质量增长最快的阶段,恢复期有1-2周,故影响了模型组小鼠的最终体质量。也说明高脂食物对小鼠存在应激反应,自由摄食时,小鼠可以通过减少摄食量逐步适应,所以在灌胃造模时,应该由小量(每10 g体质量0.15 mL剂量)在1周内逐步增加至正常量(每10 g体质量0.3 mL剂量),以减小应激反应的影响。
肝脏病理学改变是诊断NAFLD最重要的诊断依据。本实验中,模型组小鼠在第12周的肝脏病理片已经表现出大量肝细胞(超过30%)体积增大,脂质空泡,形成的典型NAFLD病理表现,16周时病理改变更加明显,加上ALT及AST水平增高,充分说明已出现肝细胞炎症坏死。血脂检查结果表明,模型组高胆固醇血症明显,LDL水平升高比HDL水平降低表现得更加迅速、明显。TG在模型组表现出下降的趋势,与刘芳等[13]研究结果相同,但是原因不明,考虑为人类与老鼠的种属差异性所致。综合上述,基础饲料联合高脂肠内营养液灌胃的方法能模拟人类NAFLD的发病过程,成功建造出稳定的中重度NAFLD模型。
NAFLD目前确切发病机制虽尚不清楚,但固有免疫细胞在引发和加重NAFLD的病理过程中起到了关键作用却是公认的[14]。除了之前研究较多的枯否细胞(Kupffer cells,KCs)、树突状细胞(dendritic cells,DCs)、中性粒细胞外,近年来对NK细胞和NKT细胞在NAFLD中的作用日益受到重视。本研究中,与对照组相比,模型组CD4+T细胞比例有下降趋势,但差异无显著性,提示辅助性T细胞在NAFLD肝脏组织中的数目无明显改变。与对照组相比,模型组肝脏NK细胞数有所增加,与肝脏ALT及AST水平改变一致,虽然差异无显著性,与Dong等[15]发现的NK细胞在肝脏募集和激活后可以引起肝损伤,出现血清中ALT、AST轻度升高的结果一致。
NKT细胞是连接先天性免疫和获得性免疫的桥梁,因能快速高效地生产Th1、Th2细胞因子,在病毒感染、肿瘤免疫、自身免疫性疾病中发挥作用[16]。在以往对NAFLD患者肝内NKT细胞的研究发现,NKT细胞的数量及IL-4的分泌量增加[17]。本研究中,模型组NKT细胞的数量及IL-4+NKT百分比均增加,与对照组相比,差异具有显著性,与NAFLD患者NKT细胞变化一致。以往的小鼠NAFLD模型研究提示,该疾病状态下,NKT细胞数量是减少的,细究其原因,不能排除造模应用了胆碱缺乏饲料[11]或者基因缺陷的模型动物[18]所导致。本研究模型的制备模拟了最接近人类疾病发生、发展过程,在NKT细胞的数量及分泌活性方面符合NAFLD的病理改变,为该疾病的研究提供了良好的动物模型,并提示NKT细胞可能是治疗该疾病的一个新靶点。
(致谢:感谢广西医科大学急诊医学实验室、广东科学院生物资源应用研究所及广州医科大学呼吸疾病国家重点实验室的老师同学对本研究的大力支持及协助!)
[1] Rinella M E. Nonalcoholic fatty liver disease: a systematic review[J].JAMA, 2015,313(22):2263-73.
[2] Goh G B, McCullough A J. Natural history of nonalcoholic fatty liver disease[J].DigDisSci, 2016,61(5):1226-33.
[3] Younossi Z M, Blissett D, Blissett R, et al. The economic and clinical burden of nonalcoholic fatty liver disease in the United States and Europe[J].Hepatology, 2016,64(5):1577-86.
[4] 罗春华, 李国静, 周 军, 等. 宜昌市职业人群非酒精性脂肪性肝病流行病学调查及其与代谢综合征相关性研究[J].重庆医学,2016,45(3):390-2.
[4] Luo C H, Li G J, Zhou J, et al. Epidemiology investigation of nonalcoholic fatty liver disease in Yichang professional crowd and its correlation with the metabolic syndrome[J].ChongqingMedJ, 2016,45(3):390-2.
[5] 潘 磊, 张金彪, 崔荣岗, 等. 非酒精性脂肪肝C57BL/6小鼠模型的建立[J].中国组织工程研究,2016,20(40):6054-9.
[5] Pan L, Zhang J B, Cui R G, et al. Establishment of nonalcoholic fatty liver C57BL/6 mouse models[J].JClinRehabilTissueEngRes,2016,20(40):6054-9.
[6] 张园园, 周希乔. 肠道菌群与非酒精性脂肪肝研究进展[J].中华消化杂志,2016,36(11):790-2.
[6] Zhang Y Y, Zhou X Q. Research progress in intestinal flora and non-alcoholic fatty liver disease[J].ChinJDig,2016,36(11):790-2.
[7] 李晓冲, 张秀英, 徐 尚, 等. C57BL/6J小鼠非酒精性脂肪肝模型的建立[J].中国兽医杂志,2013,49(1):6-8.
[7] Li X C, Zhang X Y, Xu S, et al. Establishment of models of non-alcoholic fatty liver in C57BL/6J mouse[J].ChinJVetMed,2013,49(1):6-8.
[8] 魏 伟,吴希美,李元建. 药理实验方法学[M]. 人民卫生出版社,2010:496.
[8] Wei W, Wu X M, Li Y J. Experimental Methodology of Pharmacology[M]. People’s Medical Publishing House,2010:496.
[9] Chen Y P, Zhang J H, Li C Q, et al. Obesity enhances Th2 inflammatory response via natural killer T cells in a murine model of allergic asthma[J].IntJClinExpMed, 2015,8(9):15403-12.
[10] 陈一平, 姜晓红, 封广义, 等. 不同品系小鼠肥胖哮喘模型建立及比较[J].中国药理学通报,2016,33(2):288-92.
[10] Chen Y P, Jiang X H, Feng G Y, et al. Establishment and comparison of obese asthma models in different strains of mice[J].ChinPharmacolBull,2016,33(2):288-92.
[11] Kremer M, Thomas E, Milton R J, et al. Kupffer cell and interleukin-12-dependent loss of natural killer T cells in hepatosteatosis[J].Hepatology, 2010,51(1):130-41.
[12] 王俊杰, 方会龙, 李纯伟, 等. 非酒精性脂肪肝模型小鼠的建立[J].中国组织工程研究与临床康复,2011,15(24):4395-9.
[12] Wang J J, Fang H L, Li C W, et al. Establishment of non-alcoholic fatty liver model in mice[J].JClinRehabilTissueEngRes,2011,15(24):4395-9.
[13] 刘 芳, 高南南, 杨润梅, 等. 不同品系小鼠肥胖模型比较及C57BL/6J小鼠肥胖机制研究[J].中国药理学通报,2013,29(3):360-5.
[13] Liu F, Gao N N, Yang R M, et al. Comparison of obesity models established in the different strains of mice and the mechanism of obese C57BL/6J mice[J].ChinPharmacolBull,2013,29(3):360-5.
[14] Arrese M, Cabrera D, Kalergis A M, et al. Innate immunity and inflammation in NAFLD/NASH[J].DigDisSci, 2016,61(5):1294-303.
[15] Dong Z, Wei H, Sun R, et al. Involvement of natural killer cells in PolyI:C-induced liver injury[J].JHepatol, 2004,41(6):966-73.
[16] Scheuplein F, Thariath A, Macdonald S, et al. A humanized monoclonal antibody specific for invariant natural killer T (iNKT) cells forinvivodepletion[J].PLoSOne, 2013,8(9):e76692.
[17] Tajiri K, Shimizu Y, Tsuneyama K, et al. Role of liver-infiltrating CD3+CD56+natural killer T cells in the pathogenesis of nonalcoholic fatty liver disease[J].EurJGastroenterolHepatol, 2009,21(6):673-80.
[18] Guebre-Xabier M, Yang S, Lin H Z, et al. Altered hepatic lymphocyte subpopulations in obesity-related murine fatty livers: potential mechanism for sensitization to liver damage[J].Hepatology, 2000,31(3):633-40.
猜你喜欢
航天电子对抗(2022年4期)2022-10-24
中华养生保健(2020年5期)2020-11-16
中国科技纵横(2018年2期)2018-11-29
中国生殖健康(2018年1期)2018-11-06
中国当代医药(2018年12期)2018-06-16
医药前沿(2018年8期)2018-03-15
中国比较医学杂志(2017年3期)2017-01-17
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年2期)2015-03-11
食品工业科技(2014年5期)2014-03-11
