
影响试管群落中耐药大肠杆菌比例因素的研究
2018-06-06 00:59吕世明谭艾娟刘金平张顺然
中国兽药杂志 2018年5期
杨 可,吕世明*,谭艾娟,刘金平,寇 宏,王 想,张顺然
(1.贵州大学动物科学学院,贵阳 550025;2. 贵州大学生命科学学院,贵阳 550025)
抗菌药的广泛使用,导致细菌耐药性水平不断增加[1],使动物细菌感染的控制难度加大,养殖风险和成本增高,危害养殖业的健康发展[2]。同时,动物源细菌耐药性可通过食物、饮水和接触等方式传递给人,危及人的健康[3]。如何控制耐药病原菌及其危害是摆在人类面前一个重大的公共卫生安全问题[4]。目前能有效降低细菌耐药性的措施仍旧十分匮乏,欧美一些国家采取优化用药方案[5]、限用乃至禁用抗菌药[6]等方法来减低耐药性,但效果不甚理想[7]。β-内酰胺类是目前人医和兽医临床最常用的一类抗生素[8]。2017年耐β-内酰胺类肠杆菌科细菌被WHO列入首份“用于指导新型抗生素研究和开发的全球抗生素耐药重点病原体名单”中[9]。寻求如何对抗耐β-内酰胺类抗生素的病原体已成为国际研究的热点[10]。本研究通过创建耐药菌-敏感菌,耐药菌-敏感菌-益生菌等二元和三元微生物群落[11],考察耐药菌起始比例、益生菌种类和营养水平对耐氨苄西林大肠杆菌在群落中比例变化的影响,以期为了解耐药菌生存竞争能力[12]、制定科学合理的降低和消除动物源耐药性措施提供依据。
1 材料与方法
1.1 菌株 猪源耐药大肠杆菌G414(氨苄西林MIC>512 μg/mL)由贵州大学动物药理实验室保藏;大肠杆菌标准菌株ATCC25922(批号A0246B)购自广东环凯生物科技有限公司;蜡样芽孢杆菌CICC21252、凝结芽孢杆菌CICC20138、枯草芽孢杆菌CICC20683、地衣芽孢杆菌CICC20684购自中国工业微生物菌种保藏管理中心。
1.2 药品 氨苄西林钠(批号N0521A)购自大连美仑生物技术有限公司;伊红美蓝琼脂(EMB,批号20160909)、LB肉汤(批号20161107)、米勒海顿琼脂(MHA,批号20160603)、干粉培养基由青岛海博生物技术有限公司生产。
1.3 主要仪器 SW-CT-1F超净工作台:上海博迅实业有限公司医疗设备厂;FA2004电子分析天平:上海良平仪器仪表有限公司;XH-C快速混匀器SK-1:常州澳华仪器有限公司;HH.B11-420-S-Ⅱ型恒温培养箱:上海贺德实验设备有限公司;SHZ-82气浴恒温振荡器:常州澳华仪器有限公司;LDZM-60KCS型全自动高压蒸汽灭菌锅:上海申安医疗器械厂。
1.4 试验方法
1.4.1 耐药菌比例的测定方法 参照影印培养法[13- 14]改良测定群落中耐药大肠杆菌菌株的菌落数,耐药菌氨苄西林的筛选浓度为512 μg/mL,耐药菌落比例计算公式如下:耐药菌落比例(%)=氨苄西林MHA培养基上菌落数/EMB琼脂培养基上菌落数×100%。
1.4.2 耐药大肠杆菌初始比例对耐药菌比例的影响 0.5麦氏度耐药大肠杆菌按照0、10%、33.3%的比例(V/V)加入到0.5麦氏度的标准大肠杆菌菌液中,37 ℃,培养7 d,每天离心,更换新培养基,并测定耐药菌比例。
1.4.3 益生菌种类对耐药菌比例的影响 蜡样芽孢杆菌、枯草芽孢杆菌、地衣芽孢杆菌等3种益生菌菌液制成0.5麦氏度,分别与含10%耐药大肠杆菌的0.5麦氏度标准大肠杆菌菌液等体积混合,按1.4.1项方法测定1~7 d的耐药菌比例。
1.4.4 营养水平对耐药菌比例的影响 将含10%耐药大肠杆菌的0.5麦氏度标准大肠杆菌菌液接种至相应浓度的LB肉汤,按1%低营养水平、10%中营养水平、100%高营养水平考察益生菌初始比例对耐药菌比例的影响。
以上试验组均置于恒温振荡培养箱中(37 ℃、200 r/min)培养,每隔1 d取出测量一次耐药菌比例并做更换培养液等处理,连续测量7 d。
1.5 数据处理 利用SPSS 22.0软件对测定数据进行统计和分析,采用t检验进行显著性检验。
2 结果与分析
2.1 起始比例对群落中耐药菌比例的影响 结果如图1所示。图1显示,随培养时间的延长,2种耐药菌起始比例群落中的耐药菌比例在1~7 d均逐渐上升;高初始比例组的耐药菌比例从33%上升至64%,绝对值增加31%,超过敏感菌成为群落中的优势菌,并且1~7 d的比例均高于10%组。低初始比例组第1天从10%快速上升至36%,第7天升至43%,绝对值增加33%,升高幅度极显著大于高比例组(P<0.01)。33%组对数趋势线方程(y=0.1347ln(x)+0.3306)和10%组对数趋势线方程(y=0.1318ln(x)+0.1661)斜率差异不显著(P>0.05),10%组对数趋势线方程的截距0.1661极显著地高于折线起始点0.1(P<0.01)。说明无论初始比例是高还是低,耐药菌都没有因携带耐药基因而升高其生存适应度代价,反而获得某种竞争优势,使其逐步成为群落中的优势菌;低起始比例有助于耐药菌初期的快速生长,并可获得比高起始比例组更高的绝对增长率。

图1 起始比例对耐药菌比例的影响Fig 1 Influence of initial ratios on the proportion of amp-resistant E.coli
2.2 益生菌种类对耐药菌比例的影响 结果如图2所示。与对照组相比较,枯草芽孢杆菌和地衣芽孢杆菌在第1~7天能够显著降低耐药大肠杆菌在群落中比例的升高趋势(P<0.05)。蜡样芽孢杆菌组中耐药菌比例在第1天能够降低耐药大肠杆菌在群落中比例的升高趋势,而第2天从21%陡然上升至43%,上升比例极显著地大于对照组(P<0.01),随后又大幅滑落,第五天回落到10%附近,最终降至第7天的5%,与对照组比较差异极显著(P<0.01)。对照组对数趋势线方程(y=0.1278ln(x)+0.1648)、枯草芽孢杆菌组对数趋势线方程(y=0.1257ln(x)+0.0883)、地衣芽孢杆菌对数趋势线方程(y=0.113ln(x)+0.1019)三者斜率差异不显著(P>0.05),截距差异极显著(P<0.01)。蜡样芽孢杆菌对数趋势线方程(y=-0.036ln(x)+0.2352)的斜率与其他三组相比差异极显著(P<0.01)。说明枯草芽孢杆菌和地衣芽孢杆菌只能降低耐药菌比例在各天的上升趋势且效果相近,而蜡样芽孢杆菌则具有降低耐药菌的竞争适应度、促使其消亡的能力。

图2 益生菌种类对耐药菌比例的影响Fig 2 Influence of added probiotics on the proportion of amp-resistant E.coli
2.3 营养水平对耐药菌比例的影响 结果如图3所示。图3显示,高、中、低营养条件下,1~7 d群落中耐药菌比例均逐步升高;三组的对数趋势线方程分别为y=0.1422ln(x)+0.1802(10%组)、y=0.1318ln(x)+0.1661(100%)、y=0.1125ln(x)+0.1159(1%组),1%组(低营养水平)的耐药菌比例较其他两组上升最慢,10%组(中营养水平)的上升最快,而100%组(高营养水平)的上升速度介于二者之间,三者的差异显著(P<0.05)。说明耐药菌株在一个中等营养水平环境中的相对适应度最大。
3 讨论与小结
3.1 耐氨苄西林大肠杆菌G414与标准菌株竞争力比较 细菌种内和种间的竞争是塑造试管中群落结构的重要生态过程,它与个体生长速率、繁殖率、存活率等有关[15]。经典的进化论认为,当细菌在抗菌药选择压力下耐药突变菌株数量增加,获得了在有药环境下的生存优势,而成为该环境下的优势菌群;在无药环境下,耐药菌株携带和表达耐药基因需要额外耗费细菌的资源,增加了细菌的竞争适应度代价(生存负担),降低其竞争适应度,而在与野生型细菌的竞争中处于劣势,最终被稀释,消失。很多的研究结果也证明了这个理论的正确性。但研究结果表明,除蜡样芽孢杆菌外,其他单因素水平作用下耐药菌株的生存竞争能力均强于标准菌株,说明本株耐氨苄西林大肠杆菌由于耐药突变而获得了比野生菌更强的生存能力,此结果与传统的关于耐药菌在无药环境中竞争适应度下降的理论相左。此结果与Melny[16]、Andersson[17]等报道的结果相似。
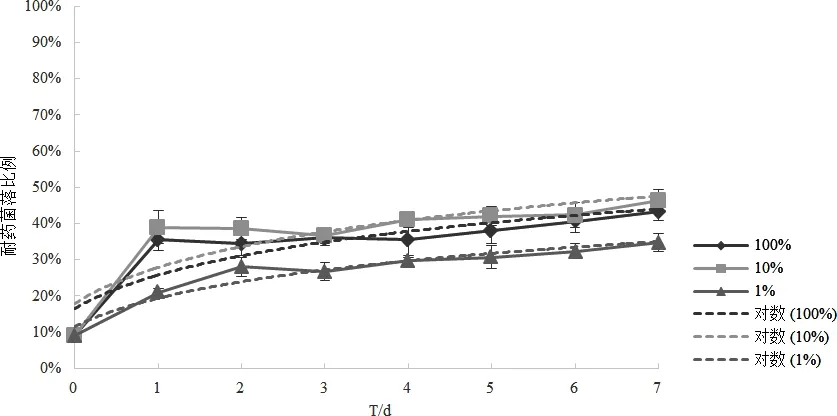
图3 营养水平对耐药菌比例的影响Fig 3 Influence of nutritional levels on the proportion of amp-resistant E.coli
3.2 益生芽孢杆菌对耐氨苄西林大肠杆菌G414生存竞争的影响 在狭小的试管中菌群间存在着营养和生存空间等资源的竞争作用,由于自然微生物群落成份过于复杂,作用机制难以弄清,可控性差,因此,人工创建的两个或多个物种共培养的微生物群落,利用微生物生态学方法,便于考察耐药大肠杆菌群落组成、功能和互作效应[18]。本研究考察了大肠杆菌耐药和标准株型之间、大肠杆菌和芽孢杆菌不同种属之间的生存竞争。研究表明所用的枯草芽孢杆菌和地衣芽孢杆菌能微弱地降低耐药菌株的生存竞争能力,而蜡样芽孢杆菌除第2天外均极显著地抑制耐药菌株的生长。这在国内外鲜有报道。若加以进一步地研究开发,用于养殖场等耐药菌密度加大的区域,可能会在一定程度上减小环境中耐药菌的比例。
3.3 耐氨苄西林大肠杆菌G414及使用氨苄西林的安全性 营养匮乏会增加物种间的生存竞争压力,加速生存淘汰。本研究表明耐药菌在高中低的营养条件下均表现出比敏感菌更强的生存竞争能力。此结果与Phan[19]等报道的结果相似。说明该耐药菌可能获得的突变机制不仅没有增加其竞争适应度代价,反而降低了其竞争适应度代价,使其在恶劣环境中获得更强的竞争优势。以此推测,此耐药菌在环境中有远高于敏感菌的生存竞争能力,很容易成为优势菌群。因此,通过停止使用药物是不能恢复耐药区域内细菌对相应药物的敏感性,这将对安全用药和人的健康构成严重威胁。应该从耐药的角度来重新评估氨苄西林的使用安全性。
研究采用人工创建的多个物种共培养的微生物群落来呈现耐氨苄西林大肠杆菌在其中的比例变化,不同于目前国内外通行的以纯培养微生物为对象的耐药性研究。在后续的研究中有必要介入一些前沿的技术手段进行微生物组方面的探索,例如用转录组从基因的表达水平研究大肠杆菌群落耐药基因动态表达和调控;用蛋白质组揭示群落中蛋白质的组成、丰度、相互关系,等等。把这些分析结果结合起来,推测其潜在的作用机制。此外,还可以构建数学模型,预测和描述耐药菌株和其他细菌的生长、互作,以便于理解其群落属性变化的基本原则,为群体水平的耐药性研究建立基础。相信从个体水平拓展到群落水平是未来微生物耐药性研究发展的一个趋势。
参考文献:
[1] Brooks B D, Brooks A E. Therapeutic strategies to combat antibiotic resistance[J]. Advanced Drug Delivery Reviews, 2014, 78: 14-27.
[2] Barriere S L. Clinical, economic and societal impact of antibiotic resistance.[J]. Expert Opinion on Pharmacotherapy, 2015, 16(2): 151-153.
[3] Huijbers P M C. Transmission of antibiotic resistance from animals to humans: Broilers as a reservoir of ESBL-producing bacteria[EB/OL]. [2017-10-11]. http://edepot.wur.nl/366347.
[4] World Health Organization. WHO's first global report on antibiotic resistance reveals serious, worldwide threat to public health[R]. 2014.
[5] Van Loon H J, Vriens M R, Fluit A C,etal. Antibiotic rotation and development of gram-negative antibiotic resistance.[J]. Am J Respir Crit Care Med, 2005, 171(5): 480-487.
[6] Brussels. Ban on antibiotics as growth promoters in animal feed enters into effect[EB/OL]. [2017-8-11]. http://europa.eu/rapid/press-release_IP-05-1687_en.pdf.
[7] Pakpour S, Jabaji S, Chénier M R. Frequency of Antibiotic Resistance in a Swine Facility 2.5Years After a Ban on Antibiotics[J]. Microbial Ecology, 2012, 63(1): 41-50.
[8] Elander R P. Industrial production of β-lactam antibiotics[J]. Applied Microbiology and Biotechnology, 2003, 61(5/6): 385-392.
[9] Tacconelli E, Magrini N,etal. Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics [DB/OL]. [2017-11-17]. http://www.who.int/entity/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1.
[10] Davies J, Davies D. Origins and evolution of antibiotic resistance[J]. Microbiology & Molecular Biology Reviews, 2010, 74(3): 9.
[11] Grosskopf T, Soyer O S. Synthetic microbial communities.[J]. Current Opinion in Microbiology, 2014, 18(4): 72.
[12] Lenski R E, Mongold J A, Sniegowski P D,etal. Evolution of competitive fitness in experimental populations ofE.coli: What makes one genotype a better competitor than another?[J]. Antonie Van Leeuwenhoek, 1998, 73(1): 35.
[13] Lederberg J, Lederberg E M. Replica plating and indirect selection of bacterial mutants.[J]. Journal of Bacteriology, 1952, 63(3): 399.
[14] Osterblad M, Leistevuo T, Huovinen P. Screening for antimicrobial resistance in fecal samples by the replica plating method.[J]. Journal of Clinical Microbiology, 1995, 33(12): 3146-3149.
[15] 李 博,陈家宽,沃金森 A. R. 植物竞争研究进展[J]. 植物学报, 1998, 15(4): 18-29.
LI Bo, CHEN JiaKuan, Andrew R Watkinson. A literature review on plant competition [J]. Chinese Bulletin of Botany, 1998, 15(4):18-29.
[16] Ah M, A W, R K. The fitness costs of antibiotic resistance mutations[J]. Evolutionary applications, 2015, 8(3): 273.
[17] Andersson D I, Hughes D. Antibiotic resistance and its cost: is it possible to reverse resistance?[J]. Nat Rev Microbiol, 2010, 8(4): 260-271.
[18] 张照婧,厉舒祯,邓 晔,等. 合成微生物群落及其生物处理应用研究新进展[J]. 应用与环境生物学报, 2015, 21(6): 981-986.
ZHANG Zhaojing,LI Shuzhen,DENG Ye,etal, Recent progress in studies of synthetic microbial community and its applications in bioengineering[J]. Chinese Journal of Applied and Environmental Biology, 2015, 21(6): 981-986.
[19] Phan K, Ferenci T. A design-constraint trade-off underpins the diversity in ecologically important traits in species Escherichia coli[J]. Isme Journal, 2013, 7(10): 2034.
猜你喜欢
电镀与精饰(2022年11期)2022-11-15
计算机仿真(2022年8期)2022-09-28
检验医学(2022年2期)2022-03-14
湖南饲料(2021年4期)2021-10-13
中西医结合心血管病电子杂志(2020年31期)2020-12-16
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
郑州大学学报(工学版)(2018年2期)2018-04-13
舰船电子工程(2010年1期)2010-04-26
