
脑梗死后癫痫与γ-氨基丁酸B受体及前强啡肽原启动子基因多态性的关系
2018-06-05 02:09秦丽微秦丽红
中国老年学杂志 2018年10期
王 璇 王 辰 秦丽微 秦丽红
(佳木斯大学附属第一医院神经内科,黑龙江 佳木斯 154002)
中国脑梗死继发癫痫发生率约为15.6%〔1〕。脑内兴奋性和抑制性神经递质失衡是癫痫发病的机制之一,其中γ-氨基丁酸、强啡肽、谷氨酸等神经递质及受体发挥关键作用〔2〕。γ-氨基丁酸B受体基因亚单位(G1465A)和前强啡肽原启动子区基因多态性与颞叶癫痫相关〔3〕,但具体调控机制及脑梗死继发癫痫易感基因方面的研究仍鲜有报道。本研究探讨脑梗死继发癫痫患者γ-氨基丁酸B受体基因(G1465A)和前强啡肽原启动子区基因多态性。
1 资料与方法
1.1一般资料 选取2015年10月至2016年10月佳木斯大学第一附属医院收治的脑梗死继发癫痫患者52例(继发癫痫组),脑梗死无癫痫患者81例(无癫痫组);诊断标准均符合急性缺血性脑卒中诊治指南(2014版),并签署知情同意书。继发癫痫组中男28例,女24例,年龄58~85岁,平均(70.61±8.37)岁,近1个月内均未服用抗癫痫药物,既往无癫痫史和其他颅内疾病。无癫痫组中男45例,女36例,年龄56~87岁,平均(69.35±10.08)岁。选取同期90例体检正常老年人为健康对照组,其中男49例,女41例,年龄60~85岁,平均(70.28±8.39)岁。三组受试者性别、年龄差异均无统计学意义(P>0.05),具有可比性。
1.2方法
1.2.1基因组DNA提取 取三组受试者空腹肘静脉血3 ml,放入抗凝剂预处理的试管中,使用全血基因组DNA小量提取试剂盒(上海索宝生物科技有限公司),柱层析法提取基因组DNA,-20℃保存待测。
1.2.2γ-氨基丁酸B受体基因(G1465A)基因型检测 采用PCR-RFLP技术检测:①PCR反应:上游引物5'-GAACGAT TGCACCCAAACTGG-3',下游引物5'-CTCGTGGCATGAGGTCCGA CC-3';反应体系30 μl(基因组DNA 0.1~0.2 μl,上游和下游引物各2 μl,2×TaqDNA聚合酶15 μl,双蒸馏水适量);常规反应条件下进行40个循环,与末次循环结束后72℃延伸5 min;取产物10 μl,20琼脂糖凝胶电泳,确定扩增成功。②RFLP检测:取PCR产物10 μl,双蒸馏水17 μl,Tango缓冲液(10×)2 μl,EagⅠ1 μl,37℃反应20 h,收集产物5 μl,琼脂糖凝胶电泳,ChemiDoc MP成像系统中观察扩增情况。③结果判定:γ-氨基丁酸B受体基因第7外显子1465位有G-A多态性,当此位为G等位基因时,则有EagⅠ酶切位点;当此位为A等位基因时,则无酶切位点。因此,PCR扩增产物为1个片段(441 bp)为A/A型,3个片段(441 bp、183 bp、258 bp)为A/G型,2个片段(183 bp、258 bp)为G/G型。
1.2.3前强啡肽原启动子区基因多态性检测 采用PCR技术检测:①PCR反应:上游引物5'-TCGTTAGTCT CCAACTTCAACCTGCG-3',下游引物5'- CGTGGTCCGCCAATCCA TCTCCAAGA-3';反应体系和反应条件同方法1.2.2。②PCR产物电泳和结果判定:取PCR产物10 μl,琼脂糖凝胶电泳,ChemiDoc MP成像系统中观察扩增情况。产物片段长度应为:590 bp、522 bp、454 bp、386 bp,含有1~4个拷贝数的重复单位(68 bp),电泳结果1个条带的为纯合子基因型,2个条带的为杂合子基因型;等位基因含1~2个拷贝数的为L型,含3~4个拷贝数的为H型。因此,前强啡肽原启动子区基因可分为:L/L型(1/1、1/2、2/2);L/H型(1/3、2/3、1/4、2/4);H/H型(3/3、3/4 、4/4)。
1.3统计学方法 应用SPSS21.0软件行χ2检验。
2 结 果
2.1γ-氨基丁酸B受体(G1465A)基因多态性检测结果 PCR扩增产物电泳显示441bp条带,见图1。37℃经EagⅠ酶切24 h后电泳仅显示258 bp和183 bp条带,见图2。提示三组受试者γ-氨基丁酸B受体基因第7外显子1465位均为G等位基因,基因型均为G/G型,无A/A型、A/G型。

1~3 继发癫痫组,4~5 无癫痫组,6~7 健康对照组;图2同图1 γ-氨基丁酸B受体基因PCR扩增产物电泳图
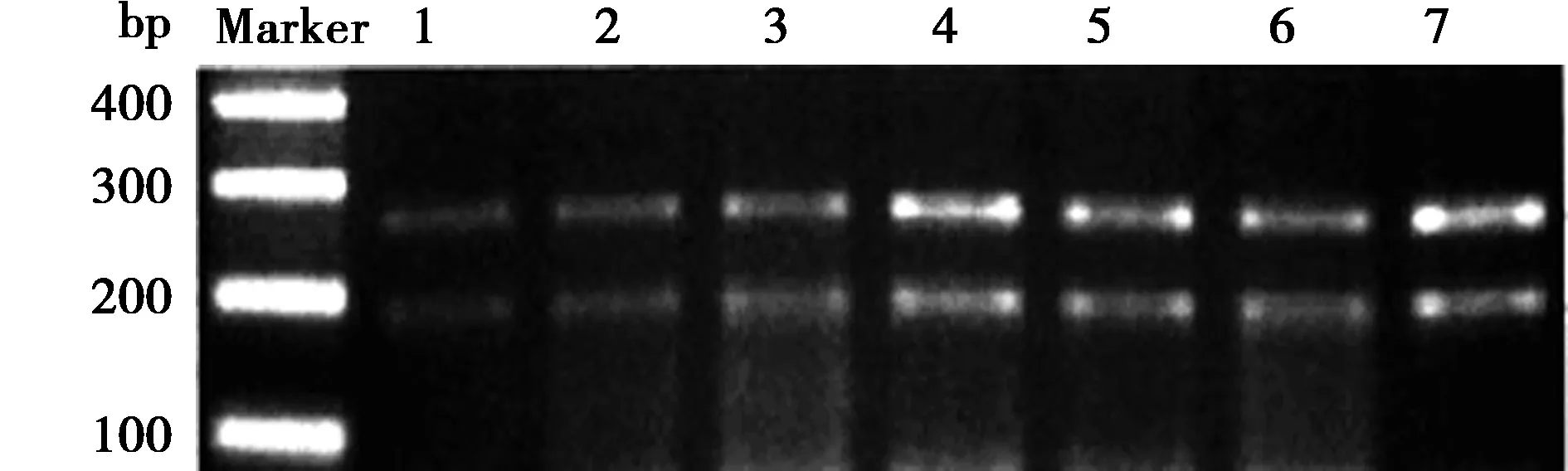
图2 γ-氨基丁酸B受体基因PCR扩增产物EagⅠ酶切电泳图
2.2前强啡肽原启动子区基因型分布情况 电泳显示,前强啡肽原启动子区基因型分别为:L/L型(1/1、2/2)、L/H型(2/3、2/4)、H/H型(3/3),见图3。继发癫痫组患者L/L型频率明显高于无癫痫组、健康对照组,H/H型频率明显低于无癫痫组、健康对照组(P<0.01);无癫痫组与健康对照组前强啡肽原启动子区基因型分布差异无统计学意义(P>0.05)。见表1。
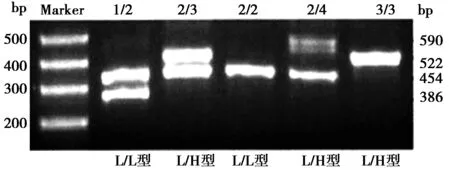
图3 前强啡肽原启动子区基因PCR扩增产物电泳图

表1 三组前强啡肽原启动子区基因型分布比较〔n(%)〕
与继发癫痫组相比:1)P<0.01
2.3前强啡肽原启动子区等位基因频率分布情况 继发癫痫组等位基因L型频率为88.46%(46/52),明显高于无癫痫组、健康对照组的14.81%(12/81)、16.67%(15/90);无癫痫组、健康对照组等位基因H型频率为85.19%(69/81)、83.33%(75/90),明显高于继发癫痫组的11.54%(6/52)(P<0.01)。无癫痫组与健康对照组前强啡肽原启动子区基因型频率差异无统计学意义(P>0.05)。
3 讨 论
研究发现,脑梗死后神经递质γ-氨基丁酸可抑制兴奋性氨基酸,减轻缺血造成的神经细胞兴奋毒性,而强啡肽原可抑制谷氨酸分泌,抑制神经细胞兴奋性,两者均具有抗惊厥作用〔4〕。已发现的γ-氨基丁酸受体包括A、B、C三种亚型,其中A、B型受体与癫痫发生、发展有明显关联〔5〕。γ-氨基丁酸受体基因变异在癫痫发病机制中的作用受到广泛关注。研究发现,γ-氨基丁酸B受体基因(G1465A)多态性与颞叶癫痫有相关性〔6〕,但之后相关研究结论与其不完全一致〔7〕;且γ-氨基丁酸B受体基因多态性是否与继发性癫痫发病相关尚未见报道。本研究显示,三组受试者γ-氨基丁酸受体基因均为G/G型,未发现A/A型、A/G型。其原因可能与人种差异有关,本次受试者均为汉族人群,而该人群中γ-氨基丁酸B受体基因第7外显子1 465位很少存在G-A突变,并非脑梗死继发癫痫的易感基因。强啡肽原为另一个抑制性神经递质,与γ-氨基丁酸具有协同作用。相关研究发现,强啡肽原主要作用于Kappa类阿片受体,且与NMDA受体结合发挥生物学作用,抑制兴奋性氨基酸诱发的惊厥〔8〕。前强啡肽是强啡肽原的前体,经酶解可生成强啡肽原。相关研究发现,人强啡肽原基因启动子区长68 bp重复单元中含有AP-1结合位点,其中AP-1复合物H型等位基因活化后可提升强啡肽原基表达水平,而相同条件下L型等位基因无法活化〔7〕。本次研究提示前强啡肽启动子区基因型对脑梗死继发癫痫有明显影响,其中L型等位基因是继发癫痫的易感基因,H型等位基因是保护基因。
4 参考文献
1黄永锋.老年性癫痫的病因及脑电图特点〔J〕.中国老年学杂志,2013;33(2):401-2.
2靳俊功.Rho激酶2在癫痫大鼠脑内表达变化及其机制的研究〔D〕.西安:第四军医大学,2014.
3Zhou CW,Ding L,Elizabeth Deel E,etal.Altered intrathalamic GABA(A)neurotransmission in a mouse model of a human genetic absence epilepsy syndrome〔J〕.Neurobiol Dis,2015;73:407-17.
4刘志广.腺苷A1受体在癫痫发病机制中的作用及其与GABA信号通路相关性的研究〔D〕.武汉:华中科技大学,2013.
5肖 奕.激动GABAC受体对大鼠癫痫发作及脑内GABA、GAT1和ρ2mRNA表达的影响〔D〕.太原:山西医科大学,2011.
6孙柳燕,臧希卉,郭 琪,等.吉林省艾滋病抗病毒治疗人群基因型耐药研究〔J〕.中国卫生工程学,2016;(3):236-9.
7赵 雪,宋文刚,兰 英,等.脂肪酶产生菌的筛选及基因的克隆表达〔J〕.北华大学学报(自然科学版),2016;17(2):186-90.
8Chowdhury FA,O'Gorman RL,Nashef L,etal.Investigation of glutamine and GABA levels in patients with idiopathic generalized epilepsy using MEGAPRESS〔J〕.J Magn Res Imag,2015;41(3):694-9.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
川北医学院学报(2022年6期)2022-06-24
昆明医科大学学报(2022年2期)2022-03-29
中国食品(2021年17期)2021-09-27
智慧健康(2021年17期)2021-07-30
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
中国动物传染病学报(2020年2期)2020-04-02
遵义医科大学学报(2020年6期)2020-02-05
中国粮油学报(2019年4期)2019-07-12
中国酿造(2017年12期)2017-12-28
