
miR-155过表达髓核细胞IncRNA表达谱芯片分析及对凋亡功能的影响
2018-06-05 02:09覃国忠史鹏亮霍少川
中国老年学杂志 2018年10期
覃国忠 史鹏亮 张 震 霍少川
(广州中医药大学,广东 广州 510000)
相关研究认为,IncRNA与miRNA存在相互调控机制,IncRNA可通过内源性吸附miRNA来调控基因表达〔1〕;同时miRNA能够绑定IncRNA,在多种RNA结合蛋白共同帮助下,诱发miRNA介导的沉默机制,调节IncRNA的稳定性〔2〕。研究已证实,miR-21与GAS5 IncRNA之间的反馈调控机制在乳腺癌、骨性关节炎等多种疾病中均有存在〔1〕。因此,人椎间盘退变中过程中miR-155能否通过IncRNA参与调控网络值得探究。本次实验先通过IncRNA芯片检测,明确miR-155过表达后椎间盘髓核细胞IncRNA表达情况,筛选细胞凋亡相关IncRNA,之后对筛选出的GAS5 IncRNA进行初步作用机制分析。
1 材料与方法
1.1材料与试剂 原代人椎间盘细胞购于美国典型微生物菌种保藏中心;hsa-miR-155过表达慢病毒载体购于上海吉凯基因化学技术有限公司;Cy3、Cy5荧光染料购于天津百萤生物科技有限公司;DMEM培养基、胎牛血清、RIPA蛋白裂解液均购于上海邦奕生物科技有限公司;含半胱氨酸的天冬氨酸蛋白水解酶(Caspase)-3抗体、B淋巴细胞瘤/白血病(Bcl)-2抗体及内参β-actin抗体购于北京百奥莱博科技有限公司。
1.2方法
1.2.1髓核细胞培养及转染 将冻存的原代人椎间盘细胞水浴复温,2 000 r/min离心10 min,2 ml髓核细胞专用培养基重悬,37℃ 5%CO2培养箱中复苏,48 h换液一次,显微镜下观察细胞融合度超过80%时用胰酶消化后加入含10%胎牛血清的DMEM培养基中传代培养。实验设研究组和阴性对照组,分别设3个平行组,标记为T1~T3和N1~N3。取对数期髓核细胞,以1×106/L接种到培养皿中,按Moi值70的病毒量转染髓核细胞,混匀后放入培养箱中孵育,6 h换液一次,每24 h在显微镜下观察细胞生长情况和转染率,转染后48 h换液,96 h在荧光显微镜下观察绿色荧光率>95%后,每个样品中滴加1 ml Trizol溶液提取细胞总RNA,低温冻存。
1.2.2miR-155过表达后髓核细胞IncRNA芯片检测 使用Agilent双通道芯片平台,将T1~T3和N1~N3互配成3对后进行芯片比较。分别用Cy3、Cy5荧光染料标记相同点阵上杂交的两个样品,标记完成经纯化处理后放入芯片扫描仪得到图像,使用相应的数据提取软件从中提取信息,记录两种荧光燃料信号值,筛选出差异性表达的IncRNA。
1.2.3筛选出的IncRNA过表达慢病毒构建及转染 筛选出的差异性IncRNA过表达慢病毒和阴性对照慢病毒由上海吉凯基因化学技术有限公司合成并测序验证。转染实验设研究组和阴性对照组,按1×106/L密度接将髓核细胞种到6孔板中,培养至细胞融合度>60%,按Moi值70的病毒量转染,混匀后放入培养箱中孵育,每24 h荧光显微镜下观察细胞生长情况和转染率,96 h后收集两组髓核细胞,提取总RNA,RT-PCR检测差异性IncRNA表达情况并进行后续实验。
1.2.4流式细胞术检测髓核细胞凋亡情况 两组髓核细胞消化离心后,以1×107/L密度重悬在190 μl结合缓冲液中,加入10 μl Annexin V-FITC,室温下避光反应15 min,2 000 r/min离心10 min,弃上清后用190 μl结合缓冲液中重悬细胞,加入10 μl PI染色液,4℃避光反应20 min,1 h内放入流式细胞仪中检测。
1.2.5Western印迹法检测Caspase-3和Bcl-2蛋白表达情况 细胞蛋白裂解液提取两组髓核细胞总蛋白,行十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳后转至聚偏氟乙烯(PVDF)膜,室温下用5%牛血清白蛋白(BSA)封闭2 h,TBST洗膜后滴加Caspase-3抗体(1∶500稀释)、Bcl-2抗体(1∶1 000稀释)、β-actin抗体(1∶1 000稀释),4℃孵育过夜,滴加Bcl-2、Caspase-3抗兔二抗和β-actin抗小鼠二抗,4℃孵育6 h,电化学发光(ECL)显色,采集图像并用电化学发光法检测。
1.3统计学分析 应用SPSS20.0软件进行t检验。
2 结 果
2.1miR-155过表达后髓核细胞IncRNA芯片分析 经IncRNA芯片分析共发现630个差异表达IncRNA,其中381个IncRNA表达上调、249个IncRNA表达下调(见图1)。差异基因筛选标准:Fc值>2倍且P<0.05。基因组数据库(Ensemble)显示,GAS5 IncRNA共包含有30个转录本,本次IncRNA芯片分析获得了13个转录本表达情况,均呈下降趋势,其中GAS5-016转录下调差异有统计学意义(Fc值≥1.5,P<0.05)。
2.2髓核细胞过表达GAS5 IncRNA后细胞凋亡情况 流式细胞术分析显示,阴性对照组髓核细胞早期凋亡率为(36.51±2.93)%,GAS5 IncRNA过表达组髓核细胞早期凋亡率为(43.75±3.34)%;GAS5 IncRNA过表达慢病毒转染72 h后髓核细胞早期凋亡率明显增加差异有统计学意义(P<0.05)。见图2。
2.3过表达GAS5 IncRNA后Caspase-3和Bcl-2蛋白表达情况 髓核细胞过表达GAS5 IncRNA后,阴性对照组和GAS5 IncRNA过表达组Caspase-3蛋白相对表达量分别为1.63±0.41、3.81±0.75;Bcl-2蛋白表相对表达量分别为7.95±0.81、2.68±0.24。GAS5 IncRNA过表达组Caspase-3蛋白表达水平明显高于阴性对照组,Bcl-2蛋白表达水平明显低于阴性对照组,差异均有统计学意义(P<0.05)。见图3。

图1 两组IncRNA表达差异聚类分析图
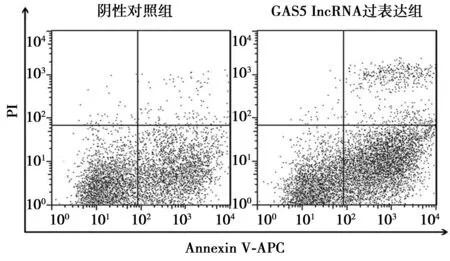
图2 髓核细胞过表达GAS5 IncRNA后细胞凋亡情况

图3 Western印迹法检测Caspase-3和Bcl-2蛋白表达
3 讨 论
本实验中采用miR-155慢病毒载体过表达人椎间盘细胞髓核细胞,使用IncRNA表达谱芯片分析显示,GAS5 IncRNA各转录本表达均出现下调,提示miR-155对人椎间盘细胞髓核细胞GAS5 IncRNA表达有负向调控作用。同时,IncRNA表达谱芯片中共检测出13个转录本表达下调,但仅GAS5-016转录本下调有显著差异,提示miR-155在人椎间盘细胞髓核细胞GAS IncRNA表达调控中,可能存在特定的一级序列特征,需进一步采用荧光素酶报告基因等检测方法探索二者可能的结合位点。
椎间盘内细胞凋亡的主要信号通路包括:内质网应激通路、线粒体途径、死亡受体通路,分别在椎间盘不同退变期发挥调控髓核细胞凋亡的作用〔3〕。线粒体在Caspase依赖性和非依赖性凋亡通路中起到重要作用,多种线粒体蛋白均可调控内源性和外源性凋亡通路〔4〕。Caspase蛋白酶和Bcl-2蛋白是哺乳动物中两个进化保守蛋白家族,其中Caspase蛋白酶可介导细胞凋亡的发生、发展〔5〕;Bcl-2蛋白可调控线粒体生理结构的完整性,抑制线粒体中cyt-c释放到胞质,进而抑制细胞凋亡〔6〕。本次研究中,GAS5 IncRNA过表达慢病毒转染72 h后髓核细胞早期凋亡率明显增加;进一步分析显示,GAS5 IncRNA过表达后明显上调Caspase-3蛋白表达,下调蛋白表达,进而激活人椎间盘细胞髓核细胞内的线粒体凋亡通路,促进髓核细胞凋亡。说明GAS5 IncRNA在髓核细胞中调控促凋亡因子和抑凋亡因子之间的平衡,其表达水平决定了髓核细胞对凋亡刺激的敏感性;同时GAS5 IncRNA过表达介导的凋亡过程中可能通过miR-155发挥作用,二者的相互调控机制仍需进一步深入研究。
4 参考文献
1张 钰,刘志礼,黄山虎,等.线粒体与椎间盘退变〔J〕.中国老年学杂志,2016;36(17):4393-6.
2陈宇飞.miR-129-5p在椎间盘退变中的作用及其表观遗传调控研究〔D〕.重庆:第四军医大学,2014.
3魏海峰,钱 军,鲍丙波,等.微血管形成在腰椎间盘退变过程中的表现特征及影响因素〔J〕.北华大学学报(自然科学版),2016;17(5):632-5.
4江 辉,王祥善.经多裂肌间隙入路微创治疗成人特发性脊柱侧凸合并椎管狭窄症临床研究〔J〕.中国卫生工程学,2017;16(1):105-7,109.
5万中元.长链非编码RNA在退变人椎间盘髓核内的异常表达〔D〕.重庆:第四军医大学,2014.
6王 华.社区康复治疗对老年骨质疏松骨折患者疼痛评分及生活质量的影响〔J〕.中国卫生工程学,2015;14(2):177-9.
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋通报(2021年1期)2021-07-23
右江民族医学院学报(2021年3期)2021-07-17
生物学通报(2021年4期)2021-03-16
中华养生保健(2020年1期)2020-11-16
保健与生活(2020年13期)2020-07-24
中国医药指南(2017年3期)2017-11-13
医学研究杂志(2015年12期)2015-06-10
癌症进展(2015年5期)2015-03-11
