
不同CO2浓度升高水平对水稻光合特性的影响
2018-05-11 08:04刘超胡正华陈健刘晓萌于凌飞孙文娟陈书涛
生态环境学报 2018年2期
刘超,胡正华*,陈健,刘晓萌 ,于凌飞,孙文娟*,陈书涛
1.南京信息工程大学气象灾害预报预警与评估协同创新中心//江苏省农业气象重点实验室,江苏 南京 210044;2.中国科学院植物研究所植被与环境变化国家重点实验室,北京 00093;3.中国科学院大学,北京 100049
随着世界工业和经济的快速发展,大气中CO2浓度逐渐增加,已由工业革命前的 280 μmol·mol-1上 升 至 当 前 的 409 μmol·mol-1( https://www.CO2.earth/daily- CO2)。在温室气体排放中等稳定化情景下,2100年大气CO2浓度预计达到538~670 μmol·mol-1(IPCC,2013)。
CO2是植物进行光合作用的重要原料,大气中CO2浓度升高必然对植物光合作用产生一定的影响(Reddy et al.,2010)。关于CO2浓度升高对植物光合作用的影响已经开展了大量研究。短期内增加CO2浓度对植物光合作用具有一定的促进作用(徐明怡等,2016)。Kitaya et al.(2004)对番茄Lycopersicon esculentum进行研究表明,CO2浓度升高为 800 μmol·mol-1条件下番茄光合速率比 400 μmol·mol-1下高1.2倍以上。王静等(2009)对小麦Triticum aestivum进行研究,结果发现在开花期和灌浆期,其净光合速率均随CO2浓度的升高而逐渐增大。然而,也有研究表明长期生长在高浓度CO2下的植物,与正常生长条件下相比,前者的光合速率接近或低于后者,即植物对高浓度CO2产生了光合适应或下调(徐明怡等,2016)。例如,廖轶等(2002)在FACE(Free-air CO2Enrichment)条件下研究水稻光合作用对CO2浓度增高的响应,发现随着处理时间的延长,高CO2浓度对水稻净光合速率的促进作用逐渐减小,且在相同CO2浓度水平下测定时,高CO2浓度处理水稻的净光合速率明显低于正常生长条件下。此外,不同植物种类以及同一种类的不同生育时期,光合速率对高CO2浓度的响应也存在很大差异(王润佳等,2010)。
CO2浓度升高对植物光合作用影响的研究多采用封闭式气室、开顶式气室(Open-top Chamber,简称OTC)或FACE试验的方法,这些研究多是建立在CO2浓度倍增或者高浓度常值基础之上(Wang et al.,2000;Singh et al.,2017),而大气 CO2浓度增加是一个渐增过程,不是瞬间增至并保持高浓度常值。瞬间 CO2浓度升高 200 μmol·mol-1以上与真实情景可能存在很大差异。因此,有必要研究不同CO2浓度升高水平(低浓度和高浓度)对植物光合特性的影响。
本研究以水稻Oryza sativa为研究对象,利用由OTC、CO2传感器、供气系统和控制系统构成的CO2浓度升高自动调控系统模拟CO2浓度增加,设置3个CO2浓度水平(以背景大气CO2浓度为对照,在背景大气CO2浓度基础上分别增加40 μmol·mol-1和 200 μmol·mol-1CO2),研究水稻光合特性对不同CO2浓度升高水平的响应,旨在指导农业生产适应气候变化,在全球气候变化大背景下对确保粮食安全具有重要意义。
1 材料与方法
1.1 田间试验
田间试验于 2016年在南京信息工程大学农业气象与生态实验站(32.16°N,118.86°E)进行。该站位于南京市浦口区盘城镇落桥村,属于亚热带湿润气候区,年平均降水量约1100 mm,多年平均温度为15.6 ℃,平均日照时数超过1900 h,无霜期为237 d。供试土壤为潴育型水稻土,灰马肝土属,耕作层土壤质地为壤质黏土,黏粒含量为26.1%。0~20 cm土壤容重为1.57 g·cm ,pH(H2O)值为6.3,有机碳、全氮含量分别为 11.95 g·kg-1、1.19 g·kg-1。
供试水稻品种为南粳9108,属常规粳稻,全生育期为149~153 d。5月20日播种,6月20日移栽。肥料运筹:氮肥施用总量为176 g·m-2,各时期氮肥施用比例为基肥∶分蘖肥∶穗肥=4∶3∶3。基肥(6月20日)采用复合肥(N∶P2O5∶K2O=1∶1∶1),分蘖期肥(7月5日)和穗肥(8月2日)均为尿素。水分及其他田间管理措施统一按当地常规栽培要求执行。
1.2 试验设计
CO2浓度设置3个水平:以背景大气CO2浓度为对照(CK),在背景大气 CO2浓度基础上分别增加 40 μmol·mol-1(T1)和 200 μmol·mol-1(T2)CO2,各处理均为4次重复,共计12个OTC。CO2浓度升高处理从水稻移栽开始至成熟结束。
不同CO2浓度升高水平由CO2自动调控系统实现(图1)。CO2自动调控系统由12个OTC、CO2传感器、供气系统和控制系统组成,实现了对OTC内CO2浓度的自动调控。
OTC为正八边形棱柱体(对边直径3.75 m,高3 m,底面积10 m2),铝合金框架,安装高透光性的普通玻璃,为聚集气体并使气体散失速率放缓,顶部开口向内倾斜45o。每个OTC内配置GMM222 CO2传感器(Vaisala Inc.,Helsinki,Finland),量程 0~2000 μmol·mol-1。
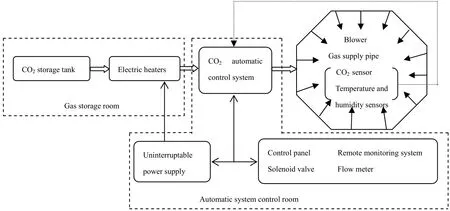
图1 CO2自动控制系统组成Fig.1 Composition of CO2 automatic control system
利用钢瓶CO2作为气源(高压液态CO2,纯度为99%),每天24 h连续供气,CO2传感器每隔2秒向自动控制系统反馈当前气室内CO2浓度。自动控制系统包括CO2感应模块、阀控制模块、流量控制模块、加压模块、主控板、数据采集等,由计算机程序实现 OTC内 CO2浓度自动监测与调节。当CO2浓度低于目标浓度时,控制系统自动打开电磁阀,实时向OTC补充CO2气体,使OTC内CO2浓度达到目标浓度。每个OTC内安装有气温和相对湿度自动监测系统,实时向控制系统传输测定数据。
1.3 测定项目与方法
1.3.1 光合-CO2响应曲线测定
利用Li-6400便携式光合作用测量系统(Li-Cor Inc.,Lincoln,NE,USA),分别于 7月 27—29日(拔节期)、8月28—29日(抽穗期)、9月23—25日(乳熟期)测定叶片净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)等相关参数。为了保证测定结果的准确性,在测定前使用设定的 1200 μmol·m-2·s-1光源诱导叶片,诱导时间不少于30 min以充分活化光合系统。测定时使用内置红蓝光源,光合有效辐射设置为 1200 μmol·m-2·s-1,叶室温度设定与实时气温保持一致,相对湿度控制在 60%~70%。采用外接 CO2小钢瓶作为气源,考虑到植物对CO2的适应性,CK处理的光合仪CO2浓度依次设定为400、300、200、100、50、400、600、800、1000、1200、1500、2000 μmol·mol-1;T1处理的光合仪 CO2浓度依次为 440、300、200、100、50、440、600、800、1000、1200、1500、2000 μmol·mol-1;T2处理的光合仪 CO2浓度依次为 600、300、200、100、50、400、440、600、800、1000、1200、1500、2000 μmol·mol-1;每个OTC选取生长状况一致的植株,叶位相同且完全展开的功能叶,在叶片中部进行测定,各处理 4个OTC观测值用于统计分析。
1.3.2 叶片全氮含量测定
将测定光合-CO2响应曲线的叶片挂上标签,光合测定完毕后,把叶片剪下,置于105 ℃烘箱中杀青30 min,随后65 ℃烘干48 h至恒重。将叶片碾磨成粉末,用元素分析仪(CHNOS Elemental Analyzer vario El III,Germany,SEAL Analytical GmbH)测定叶片全氮含量。
1.4 计算与分析
1.4.1 计算方法
叶片净光合速率与胞间 CO2浓度之间的关系用指数方程(Watling et al.,2000)进行拟合:
Pn=a(1-e-bx)+c(1)
CO2补偿点(CCP)是将胞间CO2浓度低于300 μmol·mol-1时的叶片净光合速率和 CO2浓度进行线性回归分析而得:
Pn=-c+CE×x(2)
叶片尺度水分利用效率(WUE):
WUE=Pn/Tr (3)
式(1)、式(2)、式(3)中,A为净光合速率(μmol·m-2·s-1);x为胞间 CO2浓度(μmol·mol-1);a为最大光合速率(μmol·m-2·s-1);b为羧化速率;c为呼吸速率(μmol·m-2·s-1);e为自然对数的底数;饱和 CO2浓度下净光合速率 Pnsat=a+c;羧化效率CE=b(a+c);Tr 为蒸腾速率(mmol·m-2·s-1)。当净光合速率为0时,x即为光合作用的CO2补偿点。
1.4.2 统计分析
数据采用Office Excel 2010进行预处理,运用SPSS 19.0(SPSS Inc.,Chicago,IL,USA)软件进行统计分析,各处理之间进行ANOVA分析,采用LSD法进行差异显著性检验,对总光合速率与叶片氮含量的关系进行线性回归分析。运用 Origin 2016(OriginLab Corp.,Wellesley Hills,USA)绘图软件完成制图。
2 结果与分析
2.1 净光合速率对胞间CO2浓度的响应
利用指数方程拟合各处理净光合速率(Pn)对胞间CO2浓度(Ci)变化的响应曲线(图2),并计算出光合特征参数(表1)。光合-CO2响应曲线的R2均达到了 0.96~0.99,拟合效果比较理想,能够反映各处理叶片Pn随Ci变化的趋势。
由图2可知,在拔节期、抽穗期和乳熟期,各处理Pn对Ci的响应呈现出一定的规律性变化。当Ci<400 μmol·mol-1时,Pn 随 CO2浓度增加呈近似直线上升。当 CO2浓度超过 400 μmol·mol-1时,Pn 增速逐渐降低,当 CO2到达饱和点后,Pn达到最大值后趋于稳定。在拔节期,CK实测最大净光合速率Pnmax为 39.7 μmol·m-2·s-1,T1为 40.0 μmol·m-2·s-1、T2为 40.0 μmol·m-2·s-1;在抽穗期,CK 的 Pnmax为35.0 μmol·m-2·s-1,T1为 34.2 μmol·m-2·s-1,T2为 31.4 μmol·m-2·s-1;在乳熟期,各处理 Pnmax分别为 22.6、25.0 和 20.2 μmol·m-2·s-1。同一处理不同生育时期Pnmax存在显著差异,各处理 Pnmax表现为拔节期>抽穗期>乳熟期。
在拔节期,各处理之间无显著差异。在抽穗期,当 CO2浓度在 0~200 μmol·m-2·s-1范围内时,各处理Pn随 CO2浓度的升高而增大,各处理之间差异不显著;当 CO2浓度在 200~1200 μmol·m-2·s-1范围内时,各处理间存在显著差异,Pn表现为CK>T1>T2;当 CO2浓度超过 1200 μmol·m-2·s-1后,CK 和 T1无显著差异,而T2处理较CK和T1差异显著。在乳熟期,随着 CO2浓度升高,各处理的 Pn表现为T1>CK>T2。
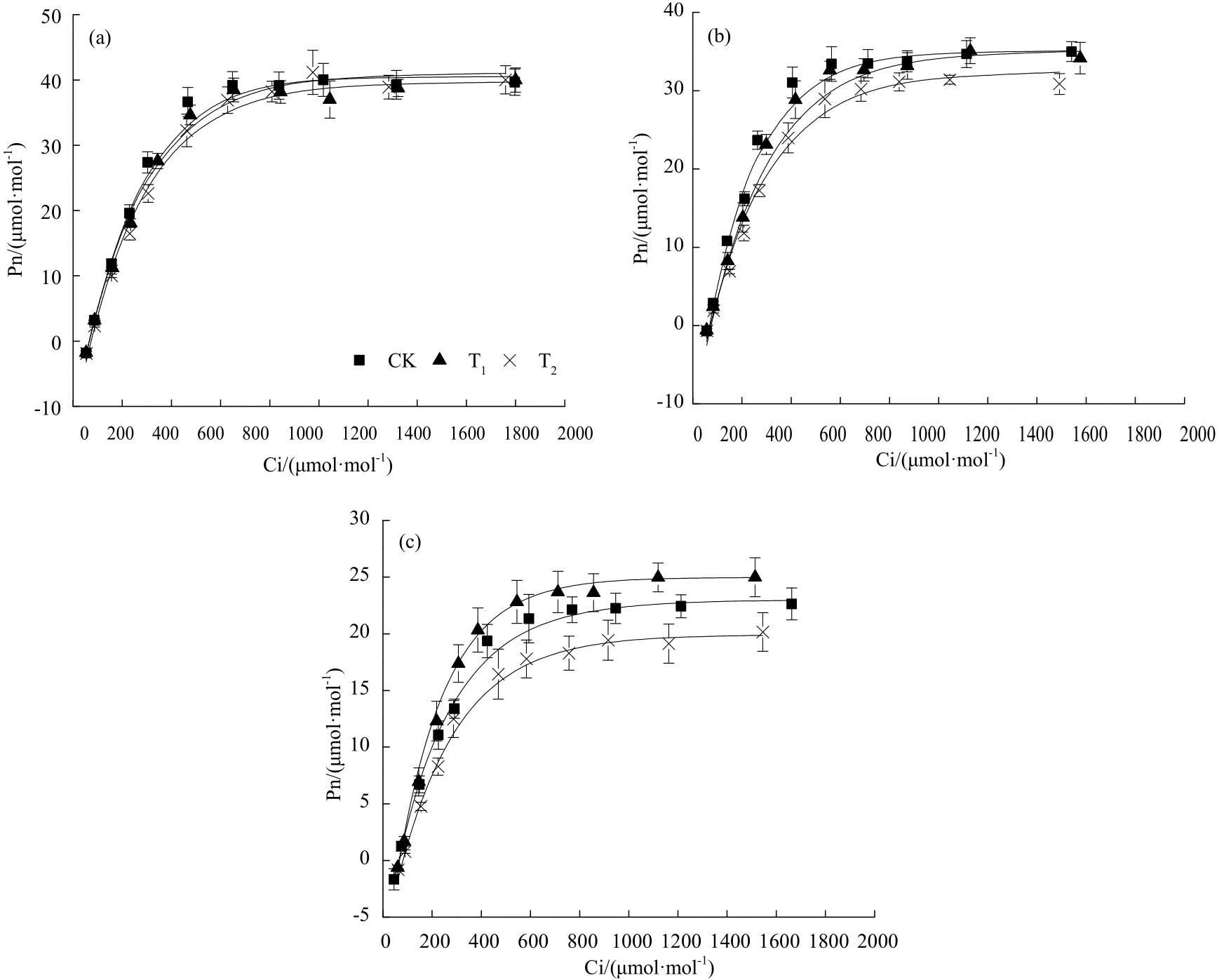
图2 净光合速率(Pn)对胞间CO2浓度(Ci)的响应Fig.2 Responses of net photosynthetic rate (Pn) to intercellular CO2 concentration (Ci)
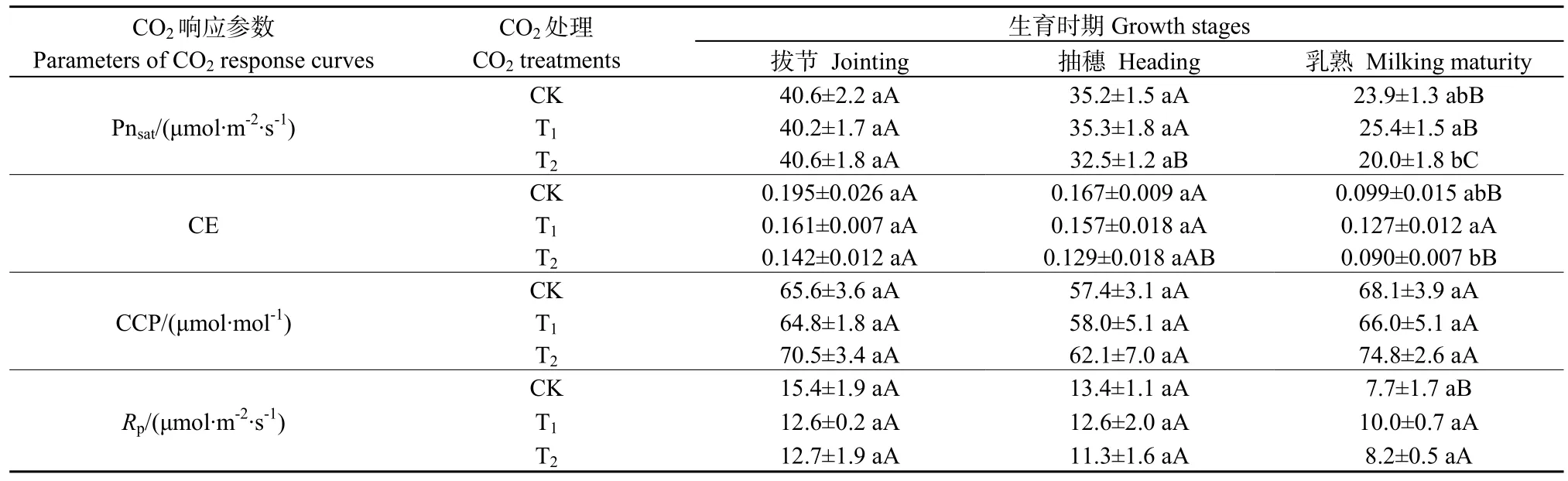
表1 不同处理下水稻不同生育时期CO2响应参数Table1 Parameters of CO2 response curves of Rice under different treatments
由表1可知,在拔节期和抽穗期,与CK相比,T1和T2处理的Pnsat、CE、CCP、Rp均不存在显著性差异;在乳熟期,T2处理下 Pnsat和CE较 T1分别显著降低了 21.3%(P<0.05)和 29.1%(P<0.05),各处理之间的CCP、Rp均不存在显著性差异。
在CK处理下,与拔节期、抽穗期相比,乳熟期Pnsat显著降低,降幅分别为41.1%(P<0.01)和32.1%(P<0.01);乳熟期CE较拔节期和抽穗期显著降低,降幅分别为 49.2%(P<0.01)和 40.7%(P<0.01);3个生育时期的CCP和Rp均不存在显著性差异。
在T1处理下,与拔节期、抽穗期相比,乳熟期Pnsat显著降低,降幅分别为36.8%(P<0.01)和28.0%(P<0.01);3个生育时期的CE、CCP和Rp均不存在显著性差异。
在T2处理下,与拔节期相比,抽穗期和乳熟期Pnsat显著降低,降幅分别为20.0%(P<0.01)和50.7%(P<0.01),乳熟期 Pnsat较抽穗期显著降低,降幅为 38.5%(P<0.01);乳熟期 CE较拔节期显著降低,降幅为36.6%(P<0.05);3个生育时期的CCP和Rp均不存在显著性差异。
2.2 CO2浓度升高对不同处理净光合速率的影响
由图 3可知,在设定 CO2浓度为 600、800、1000 μmol·mol-1条件下,CK、T1和 T2各个生育时期的 Pn表现为 Pn1000>Pn800>Pn600,说明 CO2浓度升高促进了各处理水稻叶片的 Pn。设定 CO2浓度为600 μmol·mol-1时,与CK相比,T2处理各生育时期的Pn显著降低,降幅分别为44.0%(P<0.01)、43.4%(P<0.01)和49.1%(P<0.01);不同处理各生育时期的 Pn差异均达到显著水平(P<0.05)。设定 CO2浓度为 800 μmol·mol-1时,T1和 T2处理各生育时期Pn较CK没有显著性差异,但T2处理Pn较CK分别降低了4.9%(P=0.506)、12.7%(P=0.167)和16.6%(P=0.220);CK处理乳熟期Pn较拔节期显著降低,降幅为 41.2%(P<0.01),T1和 T2处理 Pn较 CK在各生育时期均存在显著差异(P<0.05);设定 CO2浓度为 1000 μmol·mol-1时,在拔节期和抽穗期,T1和T2处理Pn较CK没有显著性差异;在乳熟期,T2处理Pn较T1显著降低,降幅为21.5%(P<0.05)。
2.3 总光合速率与叶片氮含量的关系
图4所示为各处理下总光合速率(设定CO2为600 μmol·mol-1条件)与叶片氮含量的关系。回归分析表明,总光合速率和叶片氮含量之间存在极显著正比例函数关系(P<0.01),即总光合速率随着叶片氮含量的升高而升高。CK、T1和T2处理下正比例函数的R2分别为0.81、0.67和0.61。

图 4 总光合速率(CO2=600 μmol·mol-1)与叶片氮含量的关系Fig.4 Relationship between total photosynthetic rate (CO2=600 μmol·mol-1) and leaf total nitrogen contents

图3 CO2浓度升高对不同生育时期净光合速率的影响Fig.3 Effects of elevated CO2 concentration on net photosynthetic rate of Rice in different growth stages

表2 总光合速率与叶片氮含量的关系Table2 Relationship between total photosynthetic rate and leaf nitrogen content
统计分析表明,设定CO2浓度为600、800和1000 μmol·mol-1条件下,总光合速率与叶片氮含量之间仍呈显著正相关(P<0.01)(表 2)。在回归方程中,回归系数k代表总光合速率随着叶片氮含量增加的变化速率,即叶片光合氮素利用效率(PUNE),k值越大,说明总光合速率对叶片氮含量的变化越敏感。无重复双因素方差分析表明,T2处理PUNE显著高于CK和T1,且CO2浓度越高,T2处理的PUNE越大。
2.4 叶片尺度水分利用率对胞间CO2浓度的响应
由图5可知,各处理在不同生育时期WUE均随Ci呈一定规律性变化,即WUE均随着CO2浓度的升高而增大。在拔节期,当 CO2浓度为 0~800 μmol·mol-1,各处理 WUE随 CO2浓度的升高而呈线性增加,差异不显著;当 CO2在 800~2000 μmol·mol-1范围内,各处理 WUE随 CO2浓度升高而增加的幅度逐渐减缓,T2处理WUE显著高于CK和T1。在抽穗期,各处理WUE的差异整体表现为CK>T2>T1。在乳熟期,当 CO2浓度为 0~800 μmol·mol-1时,T1处理的 WUE 高于 CK 和 T2,当CO2浓度高于 800 μmol·mol-1时,各处理之间 WUE表现为 T1>T2>CK。
CO2浓度升高对各处理叶片尺度水分利用率的影响见表3。在相同CO2浓度水平下,同一生育时期各处理WUE均不存在显著性差异。
设定 CO2浓度为 600 μmol·mol-1,与拔节期相比,CK处理抽穗期和乳熟期的WUE显著升高,升幅分别为 35.8%(P<0.05)和 42.0%(P<0.05);T1和T2处理仅乳熟期的WUE显著升高,升幅分别为55.8%(P<0.01)和41.7%(P<0.01)。
设定 CO2浓度为 800 μmol·mol-1,与拔节期相比,CK处理抽穗期和乳熟期WUE显著升高,升幅分别为 42.5%(P<0.01)、44.8%(P<0.01);T1和T2在抽穗期、乳熟期的WUE显著升高,升幅分别为35.4%(P<0.05)、136.6%(P<0.01)和37.3%(P<0.05)、42.4%(P<0.05)。
设定 CO2浓度为 1000 μmol·mol-1,与拔节期相比,CK处理抽穗期和乳熟期WUE显著升高,升幅分别为 50.3%(P<0.01)、47.6%(P<0.01);T1和T2处理在抽穗期和乳熟期的WUE显著升高,升幅分别为 42.3%(P<0.05)、143.8%(P<0.01)和38.3%(P<0.05)、42.4%(P<0.05)。
3 讨论
3.1 光合特性

图5 水分利用率对胞间CO2响应曲线Fig.5 Response of water use efficiency to intercellular CO2 concentration

表3 CO2浓度升高对不同处理水稻水分利用率的影响Table3 Effects of elevated CO2 concentration on water use efficiency of rice under different treatments
植物的光合作用是描述植物光合机构功能和运转状态的基本指标,是光合机理研究的重要内容,也是揭示光合作用调节规律的有力手段(吕扬等,2016)。植物的光合作用不仅与叶绿素含量、叶片厚度和叶片成熟程度等自身因素有关,而且还受到外界环境因子如CO2浓度、温度等的影响(叶子飘等,2008)。有关研究表明,在短时间CO2浓度增加处理下,植物的光合速率会显著增加(周玉梅等,2001),然而随着高浓度CO2处理时间的延长,这种光合速率的增强会逐渐减弱甚至消失,有的甚至出现光合作用下调现象(王为民等,2000;Chen et al.,2005)。本研究表明,CO2浓度升高不同程度促进了各处理水稻Pn(图2),这可能是因为CO2浓度的升高使其增加了对Rubisco酶结合位点的竞争性,提高了羧化效率(张淑勇等,2014);另一方面,CO2浓度的增加在某种程度上会对植物的光呼吸产生抑制作用(Drake et al.,1997),从而使Pn增加。
光合-CO2响应曲线可以反映植物光合速率随CO2浓度变化的特性,是判断植物光合能力的重要指标(罗辅燕等,2013)。选用指数方程模型对光合-CO2响应曲线进行拟合,可计算出相关光合特征参数,利用这些光合特征参数可对植物的光合生理生态特性进行评价。对植物光合生理特性反映最直接的指标是 Pn,而 Pnsat是衡量植物叶片光合能力的关键性指标(张小全等,2001;霍宏等,2007)。本研究表明,随着生育进程的推进,T2处理 Pnsat逐渐降低(表1),即T2降低了水稻最大光合潜力。CE和CCP的大小反映植物利用低浓度CO2的能力(王为民等,2000),CE越大、CCP越小,表明植物利用低浓度CO2的能力越强。本研究结果表明,T2处理CE较CK和T1显著降低了,但CCP增加了(表1),说明T2处理下水稻对低CO2浓度的利用率较低,由图2也可知,在较低CO2浓度水平下,T2处理3个生育时期Pn均较CK和T1低。
本研究还发现,当 CO2浓度分别为 600、800和 1000 μmol·mol-1时,T2处理 3 个生育时期 Pn 均较CK显著下降(图3),即产生了光合下调的现象。其可能原因是由于高CO2促进了植物的光合作用,碳水化合物在叶片内大量累积,不能够及时运输,最终产物累积造成反馈抑制或造成叶绿体的损伤(范桂枝等,2005;Sawada et al.,2001)。朱延姝等(2010)研究表明,不同品种番茄Pn在苗期、开花座果期和果实膨大期逐渐下降。本研究亦表明,随着生育时期推进,各处理水稻Pn逐渐降低。
3.2 叶片氮含量
氮素是植物合成叶绿素和各种酶的重要组成成分,其对植物各项生理活动有非常重要的作用(郑淑霞等,2007)。植物叶片中大约有 75%的氮存在于叶绿体中,且大部分被用于光合器官的构建(Takashima et al.,2004)。叶片氮含量是限制植物光合能力的关键因子之一,叶片氮含量不仅会影响叶绿素含量,而且也会对叶片内参加光合作用的酶的含量及其活性产生影响(Goedhart et al.,2010)。有关研究表明,叶片氮含量的增加有助于植物进行光合作用(尹丽等,2011)。张亚杰等(2004)研究表明,植物叶片的氮含量与光合能力存在显著正相关关系。郭盛磊等(2005)对落叶松Larix gmelinii幼苗针叶进行研究,结果发现叶片的氮含量与其Pnmax呈曲线相关,当叶片氮含量超过24 mg·g-1后Pnmax开始下降。然而,也有一些研究发现,不同物种间 Pn与叶片氮含量呈负相关或无显著相关性(Takashima et al.,2004)。本研究结果表明,不同CO2浓度下,各处理水稻叶片总光合速率与氮含量均呈极显著正相关,总光合速率随着叶片氮含量的升高而升高(图 4,表 2)。光合能力与叶片氮含量比率可以作为评估氮素投入光合效率的指标,即光合氮素利用效率(PNUE),它可以用来表示氮素利用效率与固定CO2关系的瞬时测度(Takashima et al.,2004)。实验结果表明,T2处理PNUE显著高于CK和T1,表明水稻叶片PNUE随CO2浓度增加而增加。
3.3 水分利用效率
水分利用效率是植物叶片水分利用特征的基本生理参数,是表征植物在等量水分消耗情况下,固定CO2的能力(Ogutu et al.,2013)。本研究表明,各处理WUE在3个生育时期均随着CO2浓度升高而逐渐增加(图 5),这与前人研究结果相一致(王晨光等,2015;Kim et al.,2003)。在相同CO2浓度水平下,同一生育时期不同处理间 WUE不存在显著性差异(表3)。虽然T2处理Pn在CO2浓度为 600、800 和 1000 μmol·mol-1水平下相比于CK和T1整体上均发生了下调,但是本研究发现叶片Gs随着CO2浓度的升高而逐渐降低,导致部分气孔关闭,引起Tr下降,因此T2处理WUE较CK和 T1差异不显著。有研究表明,CO2浓度从 350 μmol·mol-1升高到 700 μmol·mol-1,春小麦叶片温度升高 1~3 ℃(王润佳等,2010),温度升高可以促进叶片的蒸腾作用。本研究还发现,不同生育时期WUE整体表现为乳熟期>抽穗期>拔节期,可能的原因是不同生育时期的气温存在差别,叶片温度也会随之变化。本研究中,拔节期叶片温度约为35 ℃,抽穗期约为 32 ℃,乳熟期约为 25 ℃,最大温差近10 ℃,尽管各处理Pn随着生育时期的推进呈逐渐降低趋势,但是Tr受到叶片温度的影响(马莉等2012;伍维模等2006;),其下降速率远大于Pn,故不同生育时期WUE差异显著。
4 结论
(1)对照、增加 40 μmol·mol-1和 200 μmol·mol-1CO23种处理的水稻,随着设定CO2浓度梯度升高,Pn逐渐增加,最终达到饱和状态并趋于稳定;同一CO2浓度水平,增加 200 μmol·mol-1CO2处理的 Pn低于对照,水稻长期生长在增加200 μmol·mol-1CO2浓度环境产生了光合下调现象。同时,增加 200 μmol·mol-1CO2浓度处理降低了 Pnsat和 CE。
(2)对照、增加 40 μmol·mol-1和 200 μmol·mol-1CO23种处理,在设定不同CO2浓度下总光合速率与叶片氮含量均呈极显著正相关,总光合速率随叶片氮含量升高而升高。增加 200 μmol·mol-1CO2浓度处理下,水稻 PNUE显著高于对照和增加 40 μmol·mol-1CO2处理,且随着设定CO2浓度增加而增加。
(3)对照、增加 40 μmol·mol-1和 200 μmol·mol-1CO23种处理的水稻,随着设定CO2浓度梯度升高,WUE也逐渐增加;在同一CO2浓度水平下,各处理WUE不存在显著性差异。
致谢:南京信息工程大学王亚萍、番聪聪、王艺杰、尤新媛,中国科学院植物研究所张琳、张晨希、李昕、胡洵瑀、邓希、吕春华参加光合作用测定;中国科学院植物研究所黄耀研究员对论文的撰写和修改提出宝贵建议,一并致谢。
参考文献:
CHEN G Y, YONG Z H, LIAO Y, et al. 2005. Photosynthetic acclimation in rice leaves to free-air CO2enrichment related to both ribulose-1,5-bisphosphate carboxylation limitation and ribulose-1, 5-bisphosphate regeneration limitation [J]. Plant & Cell Physiology, 46(7): 1036.
CO2now. Earth’s home page [EB/OL]. https://www.co2.earth/daily-co2,2017-4-1/2017-4-10.
DRAKE B G, GONZALEZMELER M A, LONG S P. 1997. More efficient plants: a consequence of rising atmospheric CO2? [J]. Annual Review of Plant Physiology & Plant Molecular Biology, 48(4): 609-639.
GOEDHART C M, PATAKI D E, BILLINGS S A. 2010. Seasonal variations in plant nitrogen relations and photosynthesis along a grassland to shrub land gradient in Owens Valley, California [J]. Plant and Soil, 327(1): 213-223.
IPCC. 2013. Summary for Policymakers[C]//Climate Change 2013: The Physical Science Basis. Contributi4on of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change Cambridge: Cambridge University Press.
KIM H Y, LIEFFERING M, KOBAYASHI K, et al. 2003. Seasonal changes in the effects of elevated CO2, on rice at three levels of nitrogen supply:a free air CO2, enrichment (FACE) experiment [J]. Global Change Biology, 9(6): 826-837.
KITAYA Y, SHIBUYA T, YOSHIDA M, et al. 2004. Effects of air velocity on photosynthesis of plant canopies, under elevated CO2levels in a plant culture system [J]. Advances in Space Research, 34(7):1466-1469.
OGUTU B O, DASH J, DAWSON T P. 2013. Developing a diagnostic model for estimating terrestrial vegetation gross primary productivity using the photosynthetic quantum yield and Earth Observation data [J].Global Change Biology, 19(19): 2878-2892.
REDDY A R, RASINEN G K, RAGHAWENDRA A S. 2010. The impact of global elevated CO2concentration on photosynthesis and plant productivity [J]. Current Science, 99(1): 46-57.
SAWADA S, KUNINAKA M, WATANABE K, et al. 2001. The mechanism to suppress photosynthesis through end-product inhibition in single-rooted soybean leaves during acclimation to CO2enrichment [J].Plant & Cell Physiology, 42(10): 1093.
SINGH S K, REDDY V R, FLEISHER D H, et al. 2017. Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated CO2[J]. Photosynthetica, 55(3): 421-433.
TAKASHIMA T, HIKOSAKA K, HIROSE T. 2004. Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species [J]. Plant Cell & Environment, 27(8): 1047-1054.
WANG C R, HAN S J, LUO X B. 2000. Photosynthetic response of Pinus sylvestriformis to elevated carbon dioxide and its influential factor analysis [J]. Journal of Forestry Research, 11(3): 167-172.
WATLING J R, PRESS M C, QUICK W P. 2000. Elevated CO2induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum [J]. Plant Physiology, 123(3): 1143-1152.
范桂枝, 蔡庆生. 2005. 植物对大气 CO2浓度升高的光合适应机理[J].植物学通报, 22(4): 486-493.
郭盛磊, 阎秀峰, 白冰, 等. 2005. 供氮水平对落叶松幼苗光合作用的影响[J]. 生态学报, 25(6): 1291-1298.
霍宏, 王传宽. 2007. 冠层部位和叶龄对红松光合蒸腾特性的影响[J].应用生态学报, 18(6): 1181-1186.
廖轶, 陈根云, 张海波, 等. 2002. 水稻叶片光合作用对开放式空气CO2浓度增高 (FACE) 的响应与适应[J]. 应用生态学报, 13(10):1205-1209.
罗辅燕, 陈卫英, 陈真勇. 2013. 指数改进模型在大麦光合-CO2响应曲线中的适用性[J].植物生态学报, 37(7): 650-655.
吕扬, 刘廷玺, 闫雪, 等. 2016. 科尔沁沙丘-草甸相间地区黄柳和小叶锦鸡儿光合速率对光照强度和 CO2浓度的响应[J]. 生态学杂志,35(12): 3157-3164.
马莉, 王长荣, 王博欣, 等. 2012. 冬小麦叶片水分利用效率和光合速率研究[J]. 甘肃农大学报, 47(1): 73-77.
王晨光, 郝兴宇, 李红英, 等. 2015. CO2浓度升高对大豆光合作用和叶绿素荧光的影响[J]. 核农学报, 29(8): 1583-1588.
王静, 冯永忠, 杨改河, 等. 2009. 小麦光合特性对二氧化碳浓度升高的响应[J]. 干旱地区农业研究, 27(3): 179-183.
王润佳, 高世铭, 张绪成. 2010. 高大气CO2浓度下C3植物叶片水分利用效率升高的研究进展[J]. 干旱地区农业研究, 28(6): 190-195.
王为民, 王晨, 李春俭, 等. 2000. 大气二氧化碳浓度升高对植物生长的影响[J].西北植物学报, 20(4): 676-683.
伍维模, 董合林, 王萍, 等. 2006. 水分与氮素对南疆膜下滴灌棉花水分利用效率与蒸腾速率的影响[J]. 西北农业学报, 15(1): 11-15.
徐明怡, 倪红伟. 2016. CO2浓度升高对植物光合作用的影响[J]. 国土与自然资源研究, (2): 83-86.
叶子飘, 于强. 2008. 光合作用光响应模型的比较[J]. 植物生态学报,32(6): 1356-1361.
尹丽, 胡庭兴, 刘永安, 等. 2011. 施氮量对麻疯树幼苗生长及叶片光合特性的影响[J]. 生态学报, 31(17): 4977-4984.
张淑勇, 夏江宝, 张光灿, 等. 2014. 黄河三角洲贝壳堤岛叶底珠叶片光合作用对 CO2浓度及土壤水分的响应[J]. 生态学报, 34(8):1937-1945.
张小全, 徐德应. 2001. 18年生杉木不同部位和叶龄针叶光响应研究[J].生态学报, 21(3): 409-414.
张亚杰, 冯玉龙. 2004. 不同光强下生长的两种榕树叶片光合能力与比叶重、氮含量及分配的关系[J]. 植物生理与分子生物学学报, 30(3):269-276.
郑淑霞, 上官周平. 2007. 不同功能型植物光合特性及其与叶氮含量、比叶重的关系[J]. 生态学报, 27(1): 171-181.
周玉梅, 韩士杰, 张军辉, 等. 2001. CO2含量升高对水曲柳幼苗净光合与水分利用的影响[J]. 东北林业大学学报, 29(6): 29-31.
朱延姝, 樊金娟, 冯辉. 2010. 弱光胁迫对不同生育期番茄光合特性的影响[J]. 应用生态学报, 21(12): 3141-3146.
猜你喜欢
今日农业(2021年6期)2021-11-27
内蒙古民族大学学报(自然科学版)(2021年4期)2021-09-02
中国种业(2021年2期)2021-03-01
草地学报(2019年3期)2019-07-25
中国奶牛(2019年1期)2019-02-15
新课程·下旬(2017年7期)2017-08-14
动物营养学报(2015年10期)2015-12-01
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年2期)2011-10-26
