
科尔沁沙地恢复过程中地上定植群落与土壤种子库特征及其关系研究
2018-05-11 08:04赵丽娅李元哲陈红兵张劲张晓雨
生态环境学报 2018年2期
赵丽娅,李元哲,陈红兵,张劲,张晓雨
湖北大学资源环境学院,湖北 武汉 430062
科尔沁沙地地处松嫩草原与内蒙古高原的过渡地带,由于滥垦、滥牧等导致植被被大量破坏,土地风蚀、沙化严重,生态环境脆弱,是中国北方农牧交错地区典型的生态脆弱带。该沙区降水量在350~450 mm之间,自然条件较为优越,本底植被为温带疏林草原,大部分沙地主要由草原开垦转化而来。如果停止开垦、滥牧,沙地就有逆转恢复的可能(赵学勇等,2009;段翰晨等,2013)。沙地植被恢复不仅是地上植物群落结构与组成的变化,而且土壤种子库组成与大小也将发生变化,两者是相互关联的。一方面,地上植被生产的植物种子是土壤种子库的主要来源,地上植物的生长发育状况及繁殖能力直接影响着种子库的组成及动态变化;另一方面,土壤种子库作为重要的种源储备库,通过种子的萌发及形成的幼苗参与地上植被的自然更新,直接影响着地上植物群落的组成与结构模式(李锋瑞等,2003;Kim et al.,2005)。本文以科尔沁沙地群落为研究对象,将地上植物群落与土壤种子库这两个过程有机地联系起来进行研究,以期为退化植被的恢复与重建寻求有效的途径与方法。
1 自然条件和研究方法
1.1 研究区自然条件
研究区位于科尔沁沙地中南部的奈曼旗中国科学院奈曼沙漠化研究站境内。地理位置120°41′E,42°54′N,海拔高度为 375~395 m。该区属温带半干旱大陆性气候,年平均降水量366 mm,年蒸发量1935 mm,年均气温6.5 ℃,1月平均气温-13.1 ℃,7月平均气温23.7 ℃,≥10 ℃积温在3000 ℃以上,无霜期 150 d。土壤类型为沙质栗钙土,经破坏后退化为流动风沙土,沙土基质分布广泛,风沙活动强烈。受沙土干燥疏松和研究区干旱多风等的影响,该地区在过度放牧、过垦等人为不合理利用的情况下,植被遭受破坏,土地沙化。2003年,奈曼旗颁布了《奈曼旗禁垦禁牧暂行办法》,实施禁止放牧、禁止开垦等手段,采取建设农田防护林、防风固沙林、封山育林和飞播造林种草等多种恢复措施,同时沙土有良好的水分储藏性能,在大量天然种源存在的情况下,可以使植被快速建立并向稳定植被的类型演替。
固定沙地:流沙面积低于10%,地表覆盖固定风沙土或沙质栗钙土或已经形成较厚的土壤结皮,植被盖度大于 50%,主要植物种有达乌里胡枝子(Lespedeza davurica)及一年生的猪毛菜(Salsola collina)、狗尾草(Setaria viridis)、画眉草(Eragrostis pilosa)等。
半流动沙地:流沙面积一般大于50%,地表30%覆盖物理结皮和少量生物结皮,植被盖度一般小于31%,主要植物种是大果虫实(Corispermum macrocarpum)、狗尾草。
流动沙地:地表完全被流沙覆盖,无结皮或少量物理性结皮,植被盖度少于15%,主要物种有大果虫实、沙米(Agriophyllum squarrosum)及分散的差巴嘎蒿(Artemisia halodendron)和狗尾草。
1.2 土壤种子库测定与植被调查
在禁牧封育(禁牧10年)的缓起伏沙地上,选择具有典型代表性的流动沙地(S)、半流动沙地(SS)和固定沙地(F)来表征退化沙地的3个恢复阶段,每种样地类型选取2个,分别在流动、半流动和固定沙地上设置固定观测点,在每个样地的典型地段等距离(20 m,南北向)设置 3条长150 m的样带,在每条样带上等距离(10 m)设置15个固定样点,每个样地布设45个样点,3次重复,共计270个样点,用于土壤种子库的测定和植被调查。
2013年4月初,用专门设计制作的土壤种子采样器在每个样点采集深度为5 cm、面积为20 cm×20 cm的原状土体,装入布袋,带回试验室,将土样过筛除去杂物后,均匀平摊在发芽盆内(盆底部钻有输水孔),厚度约为2 cm,然后置于玻璃温室中进行种子发芽试验和幼苗种属鉴定。种子发芽期间,每天定时喷洒适量水分,使盆内的土壤保持湿润状态,以保证尽可能多的植物种子萌发出苗。种子萌发出苗后,仔细观察和鉴定幼苗种属,一旦能够判别出一棵幼苗的种属,记其数(视其为有生命力的种子)并将其从盆中轻轻拔掉,直到识别出所有幼苗的种属。土壤种子库密度用单位面积(1 m2)土壤中有生命力的种子数量(即有效种子数量)表示。
植被调查在每个土壤种子库采样点旁边设置面积为1 m2的固定样方,8月中下旬调查每个样方的植物种组成、总盖度及每种植物的分盖度、株高与密度。并采用收获法测定地上生物量,烘箱内75~80 ℃温度下烘干48 h后测定干重。各样地每种植物的出现频率根据45个样点的测定数据计算。
1.3 数据处理方法
采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较沙地自然恢复进程中土壤种子库、地上定植群落的总密度以及每种植物密度的差异;采用线性和非线性回归分析确定土壤种子库与地上植被之间的关系。进行分析之前,对土壤种子库密度和地上植被密度数据进行对数转换。以上分析均在SPSS 18.0 for windows软件中完成。
采用 Sorensen相似性系数(similarity coefficient,SC)表征退化沙质草地土壤种子库与地上植被组成的相似性,计算公式如下:
SC=2w/(a+b) (1)
式中,SC是相似性系数;w为土壤种子库和地上植被共有的植物种数;a和b分别为土壤种子库和地上植被的植物种数(Arroyo et al.,1999)。
有关物种多样性的测度方法有很多,本研究群落物种多样性采用被广泛应用的Simpson指数(D)和Shannon-Wiener指数(H)进行分析,并在此基础上进行Pielow均匀度指数(E)的计算。
Simpson指数:

式中,S为物种总数;Pi为第i种植物的种数占群落中总种数的比例。鉴于群落中物种的其他信息,如盖度、生物量、重要值等指标更能反映群落的本质特征,个体数可由这些指标代替。本文用重要值代替个体数进行计算。
种子期和幼苗期的重要值:

上生物量;D、C、F和B分别是每一采样点调查样方中的总平均密度、总平均盖度、总样方数和总平均地上生物量。
2 结果与分析
2.1 种子库组成与密度
方差分析结果表明(表1),沙地恢复过程中,流动沙地与固定沙地土壤种子库密度存在显著差异。与流动沙地相比,固定沙地土壤种子库密度植物种数和密度分别增加了33%和95%。不同植物种的土壤种子库密度变化特征也明显不同。差巴嘎蒿的土壤种子库在半流动沙地最大,流动沙地次之,固定沙地最小,糙隐子草(Cleistogenes squarrosa)、虎尾草(Chloris virgata)、画眉草、三芒草(Aristida adscensionis)和狗尾随恢复进程的推进呈现增加趋势,沙米则呈逐渐下降趋势。
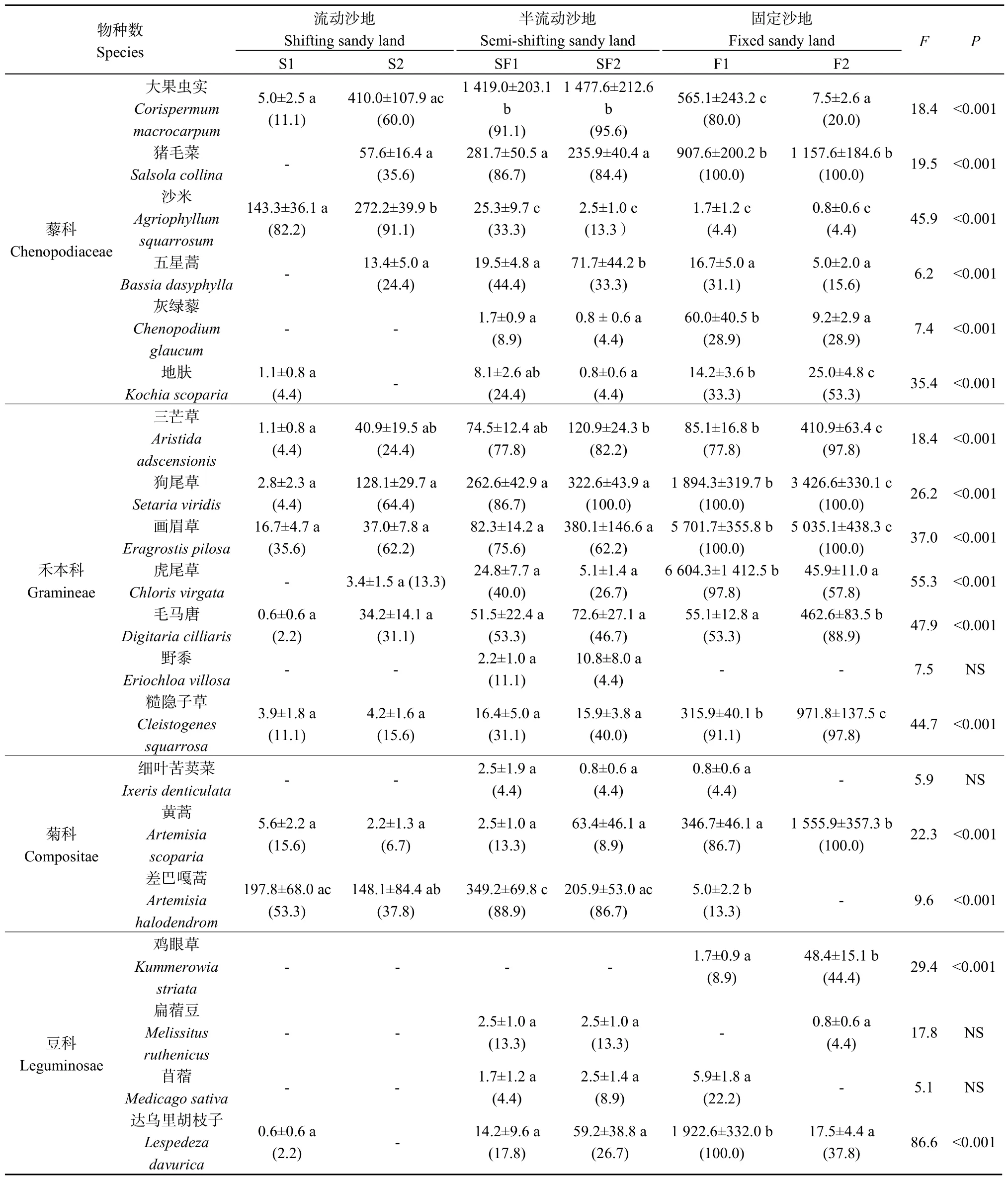
表1 土壤种子库组成、密度(ind·m-2)及频度变化特征Table1 Species composition, frequency and density (ind·m-2) of the soil seed bank in sandy lands along natural restoration gradients

续表1Continued Table1
土壤种子库群落均以一年生植物为主。流动沙地(S1)一年生植物占88%,最丰富的植物是差巴嘎蒿和沙米,其种子库密度占土壤种子库总密度的84%;流动沙地(S2)一年生植物占73%,最丰富的植物是大果虫实(Corispermum macrocarpum)、沙米、差巴嘎蒿和狗尾草,占78%的比例。半流动沙地(SS1)一年生植物占 78%,最丰富的植物是大果虫实、差巴嘎蒿、猪毛菜和狗尾草,其种子库密度占种子库总密度的 81%;半流动沙地(SS2)一年生植物占78%,最丰富的植物是大果虫实、画眉草、狗尾草、猪毛菜、差巴嘎蒿,占77%的比例。固定沙地(F1)一年生植物占83%,最丰富的植物是虎尾草、画眉草、大果虫实、狗尾草、达乌里胡枝子和猪毛菜,其种子库密度占种子库总密度的89%;固定沙地(F2)一年生植物占78%,最丰富的植物是画眉草、狗尾草、黄蒿(Artemisia scoparia)、猪毛菜和糙隐子草,占90%的比例。
2.2 地上定植群落组成与密度
由表 2可知,沙地恢复过程中,流动沙地与固定沙地地上定植群落密度存在显著差异。与流动沙地相比,固定沙地地上定植群落植物种数和密度分别增加了65%和90%。不同植物的地上定植群落密度变化也不同:沙米退出固定沙地群落;狗尾草、画眉草、虎尾草、三芒草、毛马唐(Digitaria cilliaris)、糙隐子草的密度在固定沙地中最大,半流动沙地次之,流动沙地最小;差巴嘎蒿的密度在半流动沙地中最大,流动沙地次之,固定沙地最小。
地上定植群落也以一年生植物为主。流动沙地(S1)一年生植物占 67%,最丰富的植物是沙米,其密度占地上定植群落总密度的 97%;流动沙地(S2)一年生植物占 67%,最丰富的植物是大果虫实和狗尾草,占89%的比例。半流动沙地(SS1)一年生植物占50%,最丰富的植物是大果虫实、猪毛菜和狗尾草,其密度占地上定植群落总密度的87%;半流动沙地(SS2)一年生植物占 55%,最丰富的植物是大果虫实、猪毛菜、狗尾草、三芒草和画眉草,占82%的比例。固定沙地(F1)一年生植物占68%,最丰富的植物是虎尾草、猪毛菜、狗尾草、画眉草和大果虫实,其密度占地上定植群落总密度的84%;固定沙地(F2)一年生植物占69%,最丰富的植物是画眉草、猪毛菜、狗尾草、糙隐子草和毛马唐,占91%的比例。

表2 地上定植群落组成、密度(plant∙m )和频度变化特征Table2 Species composition,frequency and density (plants·m-2) of the final established community in sandy lands along natural restoration gradients
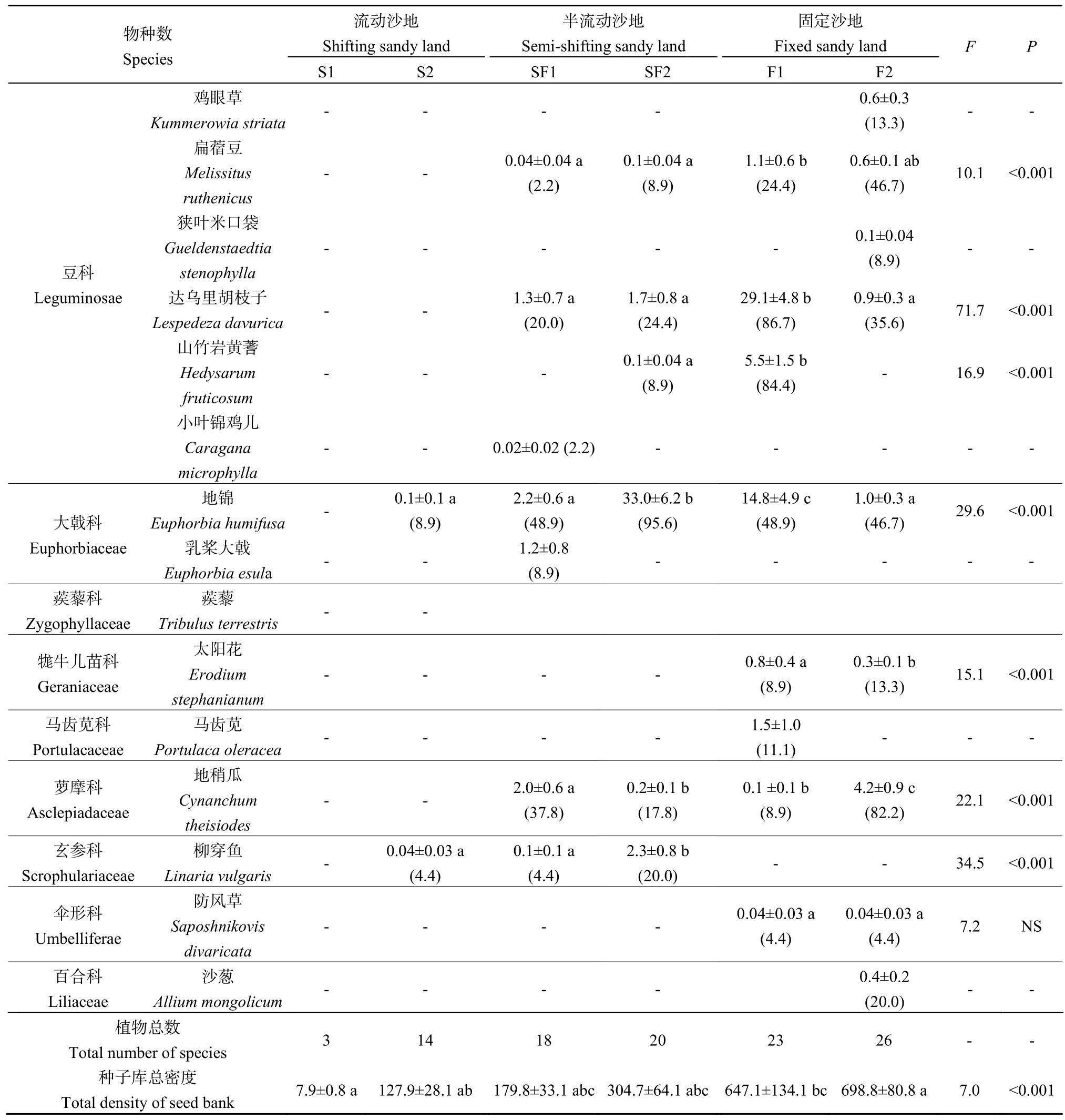
续表2Continued Table2
2.3 地上植被和土壤种子库的多样性指数
选取Shannon-Wiener指数、Simpson指数、均匀度指数和丰富度指数测度物种多样性(见表3)。结果表明,不管是土壤种子库,还是地上定植群落,物种多样性指数均表现为:固定沙地>半流动沙地>流动沙地,这与土壤种子库密度的大小顺序一致。
2.4 地上植被与土壤种子库的关系
由Sorensen相似性系数可知(表4),地上植被与土壤种子库组成的相似性系数为0.333~0.739,固定沙地的相似性系数最大,流动沙地的最小,说明土地沙漠化对地上植被组成的影响大于对土壤种子库组成的影响。
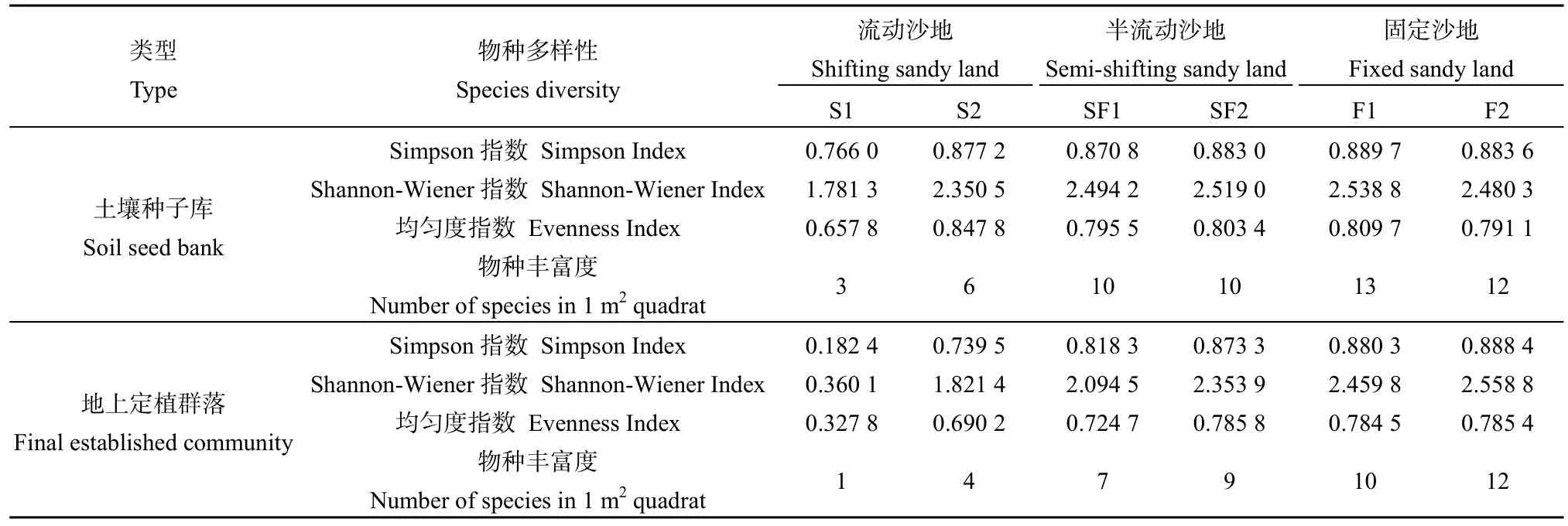
表3 土壤种子库与地上定植群落的物种多样性指标Table3 Species diversity of soil seed bank and final established community of sandy lands along natural restoration gradients

表4 地上植被与土壤种子库种类组成上的相似性系数Table4 Sorensen's similarity coefficient (SC) of species composition in soil seed bank and final established community of sandy lands along natural restoration gradients
回归分析结果显示(图 1),沙地地上定植群落与土壤种子库密度存在显著的正相关关系。该结果表明,随着土壤种子库密度的增加,地上植被密度随之增加,流动沙地、半流动沙地、固定沙地地上植被密度分别有 36%、52%、60%的变异可归结为土壤种子库密度的变异。
3 讨论和结论
3.1 讨论
土壤种子库种类组成和密度在不同恢复阶段差异显著。科尔沁沙地自然恢复过程中,流动沙地、半流动沙地、固定沙地土壤种子库平均密度分别为814、3125和16294 ind·m-2。大部分森林土壤种子库密度为102~103ind·m-2,草地土壤种子库密度为102~106ind·m-2,干旱荒漠的土壤种子库密度为 10~102ind·m-2(Marone et al.,1998;李秋艳等,2005;李宁等,2006;马建兵等;2009;唐毅等,2012;黄振英,2012)。内蒙古克氏针茅(Stipa krylovii)草原土壤种子库在6245~9156 ind·m-2之间(李元恒等,2014),西班牙中部半干旱石膏生态系统的土壤种子库规模平均为16214 ind·m-2(Caballero et al.,2003),内蒙古短花针茅(Stipa breviflora)荒漠草原的土壤种子库密度在 2554~19533 ind·m-2之间,本研究与其他半干旱地区草地的土壤种子库处于同一量级(闫瑞瑞等,2011);与库布齐沙漠、锡林郭勒退化草原不同恢复阶段的土壤种子库密度变化规律一致(于洁等,2015;仝川等,2008)。
科尔沁沙地自然恢复过程中,土壤种子库物种组成较为简单,均以一年生草本植物为主,流动沙地、半流动沙地和固定沙地的一年生植物分别占81%、78%和81%,这也是全球荒漠区常见的一个特点(李宁等,2006;孙建华等,2005;张涛等,2006;李雪华等,2006)。随着沙地的恢复,不同沙地的优势物种会出现更迭,从流动沙地到固定沙地,沙生植物沙米退出固定沙地,画眉草、虎尾草、黄蒿、糙隐子草和达乌里胡枝子等进驻群落,群落物种组成多样化、结构趋于复杂,群落的稳定性提高。
科尔沁沙地恢复过程中,地上定植群落的物种组成与密度变化与土壤种子库的变化保持一致,充分说明土壤种子库中的植物种子通过萌发和形成实生苗参与地上植被的自然更新,直接影响着地上植物群落的组成与结构(李锋瑞等,2003;李秋艳等,2005;Kassahun et al.,2009;Liu et al.,2009;Kalamees et al.,2012)。反之,地上植物的生长发育状况及繁殖能力直接影响着土壤种子库的组成、大小和动态,种子库也直接影响地上植物群落结构、组成及物种多样性(Meissner et al.,1999;Fenner et al.,2005)。此外,土壤种子库中的物种数高于地上植被,说明种子库群落组成的稳定性高于地上植被(王国栋等,2013;刘庆艳等,2014)。

图1 地上定植群落与土壤种子库密度的关系Fig.1 Relationship between established community density and soil seed bank density
土壤种子库和地上植被的关系主要有两种情况,具有相似性、相关性和不具相似性和相关性。科尔沁沙地地上植被与土壤种子库密度存在显著的正相关关系科尔沁沙地地上植被与土壤种子库在组成上的相似性系数在 0.333~0.739之间,支持以一年生草本植物为主的草原植物群落,土壤种子库与地上植被的相似性比较大,这是因为一年生植物繁殖周期短、种子产量大,以有性繁殖为主(Maranon,1998)。相反,以多年生草本植物占优势的草原植物群落,土壤种子库与地上植被的相似性很小,是因为多年生草本植物一般具有较低的种子产量,或以营养繁殖为主。例如岷江干旱河谷区灌丛植被土壤种子库和地上植被的组成物种均多为多年生植物,地上灌丛与其土壤种子库的密度及物种数均呈不显著相关性,物种组成的相似性指数较低,不足30%(李彦娇等,2010)。此外,常受干扰的生境,其种子库组成与地上植被组成相似性较高。但随着群落的成熟和干扰的减少,两者的区别变大。如森林生态系统演替早期种子库与地上植被有很大的相似性,随着林龄的增加和外来种的入侵,相似性会降低(Arroyo et al.,1999;唐勇等,1999;Olano et al.,2002;赵凌平等,2012)。例如Wanger et al.(2003)对法国Soomaa自然公园的半人工草地进行研究,发现土壤种子库与地上植被的相似性随着弃耕时间的延长而降低。因此,种子库与地上植被间的关系很难有统一的定论。不同植物在自然环境或干扰环境中的生活史对策不同,繁殖策略、种子产量、种子大小、分布空间、环境等都可影响某一物种对地上植被和种子库的贡献。或者地上植被没有进入种子库,或者与种子库的取样方法等有关。
退化草地生态系统的恢复是否可以完全依赖土壤种子库?通过对不同退化程度的草甸土壤种子库和现存植被组成的比较研究,发现在不引进原物种种子的情况下,该草甸自然恢复只能在轻度退化地段发生(Matus et al.,2003)。刘美珍(2004)研究表明,浑善达克沙地种子库是植被自然恢复的重要物质基础。土壤种子库作为一种自然力在植被恢复中起着重要作用,任何生境都可以利用土壤种子库进行原有植被的恢复。然而,对于干扰严重的环境,还要辅以人工措施逐步达到原有植被的恢复。科尔沁沙地自然本底较好,土壤种子库中含有大量的种子,在植被自然恢复中表现出极大的潜力,并能发挥重要的作用。
3.2 结论
(1)科尔沁沙地恢复过程中,流动沙地、半流动沙地、固定沙地土壤种子库的平均密度分别为814、3125和16294 ind·m-2,地上定植群落与土壤种子库的变化趋势一致,平均密度分别为 68、242和673 plants·m-2。土壤种子库和地上定植群落物种组成以一年生草本为主,多年生植物和灌木类所占的比例较小。
(2)不管是土壤种子库还是地上定植群落,Shannon-Wiener指数、Simpson指数、均匀度指数和丰富度指数的大小顺序均为:固定沙地>半固定沙地>流动沙地,土壤种子库物种多样性指数均大于地上植被。
(3)科尔沁沙地地上植被与土壤种子库组成上的相似性系数在0.333~0.739之间。回归分析表明,地上植定植群落与土壤种子库密度存在显著的正相关关系。
参考文献:
ARROYO M T K, LOHENGRIN A, CASTOR C C. 1999. Persistent soil seed bank and standing vegetation at a high alpine site in the central Chilean Andes [J]. Oecologia, 119(1): 126-132.
CABALLERO I, OLANO J M, LOIDI J, et al. 2003. Seed bank structure along a semi-arid gypsum gradient in Central Spain [J]. Journal of Arid Environment, 55(2): 287-299.
FENNER M, THOMPSON K. 2005. The Ecology of Seeds [M].Cambridge: Cambridge University Press.
KALAMEES R, PSSA K, ZOBEL K, et al. 2012. Restoration potential of the persistent soil seed bank in successional calcareous (alvar)grasslands in Estonia [J]. Applied Vegetation Science, 15(2): 208-218.
KASSAHUN A, SNYMAN H A, SMIT C N. 2009. Soil seed bank evaluation along a degradation gradient in arid rangelands of the Somli region, eastern Ethiopia [J]. Agriculture, Ecosystems and Environment,129(4): 428-436.
KIM K D, LEE E J. 2005. Soil seed bank of the waste landfills in South Korea [J]. Plant and Soil, 271(1-2): 109-121.
LIU M Z, JING G M, YU S L, et al. 2009. The role of soil seed banks in natural restoration of the degraded Hunshandak sandlands, Northern China [J]. Restoration Ecology, 17(1): 127-136.
MARANON T. 1998. Soil seed bank and community dynamics in an annual-dominated Mediterranean salt-marsh [J]. Journal of Vegetation Science, 9(3): 371-378.
MARONE ROSSI B E, CASENAVE J L. 1998. Granivore impaction soil-seed reserves in the central Monte desert, Argentina [J]. Functional Ecology, 12(4): 640-645.
MATUS G, VERHAGEN R, BEKKER R M, et al. 2003. Restoration of the Cirsio dissecti-Molinietum in The Netherlands:Can we rely on the soil seed banks? [J]. Applied Vegetation Science, 6(1): 73-84.
MEISSNER R A, FACELLI J M. 1999. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrub lands of South Australia [J]. Journal of Arid Environment, 42(2): 117-128.
OLANO J M, CABALLERO I, LASKURAIN N A, et al. 2002. Seed bank spatial pattern in a temperate secondary forest [J]. Journal of Vegetation Science, 13(6): 775-784.
WANGER M, POSHLOD P, SETCHFIELD R P. 2003. Soil seed bank in nanaged and abandoned semi-natural meadows in Soomaa national park, Estonia [J]. Annales Botanici Fennici, 40(2): 87-100.
段翰晨, 王涛, 薛娴, 等. 2013. 基于RS与GIS的科尔沁沙地沙漠化时空演变[J]. 中国沙漠, 33(2): 470-477.
黄振英, 曹敏, 刘志民, 等. 2012. 种子生态学: 种子在群落中的作用[J].植物生态学报, 36(8): 705-707.
李锋瑞, 赵丽娅, 王树芳, 等. 2003. 封育对退化沙质草地土壤种子库与地上群落结构的影响[J]. 草业学报, 12(4): 90-99.
李宁, 冯固, 田长彦. 2006. 塔克拉玛干沙漠北缘土壤种子库特征及动态[J]. 中国科学D辑, 36(增刊Ⅱ): 110-118.
李秋艳, 赵文智. 2005. 干旱区土壤种子库的研究进展[J]. 地球科学进展, 20(3): 350-358.
李雪华, 李晓兰, 蒋德明, 等. 2006. 干旱半干旱荒漠地区一年生植物研究综述[J]. 生态学杂志, 25(7): 851-856.
李彦娇, 包维楷, 吴福忠. 2010. 岷江干旱河谷灌丛土壤种子库及其自然更新潜力评估[J]. 生态学报, 30(2): 399-407.
李元恒, 韩国栋, 王正文, 等. 2014. 内蒙古克氏针茅草原土壤种子库对刈割和放牧干扰的响应[J]. 生态学杂志, 33(1): 1-9.
刘美珍. 2004. 浑善达克退化生态系统恢复研究——自然力在沙地草地恢复中的应用[D]. 北京: 中国科学院研究生院(植物研究所).
刘庆艳, 王国栋, 姜明, 等. 2014. 三江平原沟渠土壤种子库特征及其与地上植被的关系[J]. 植物生态学报, 38(1): 17-26.
马建兵, 张希明, 闫海龙, 等. 2009. 塔克拉玛干沙漠土壤种子库特征初探[J]. 干旱区研究, 26(1): 71-77.
孙建华, 王彦荣, 曾彦军. 2005. 封育和放牧条件下退化荒漠草地土壤种子库特征[J]. 西北植物学报, 25(10): 2035-2042.
唐毅, 刘志民. 2012. 沙丘生态系统种子库现状、趋势与挑战[J]. 植物生态学报, 36(8): 891-898.
唐勇, 曹敏, 张建侯, 等. 1999. 西双版纳热带森林土壤种子库与地上植被的关系[J]. 应用生态学报, 10(3): 279-282.
仝川, 冯秀, 张远鸣, 仲延凯. 2008. 锡林郭勒退化草原不同禁牧恢复演替阶段土壤种子库比较[J]. 生态学报, 28(5): 1991-2001.
王国栋, MIDDLETON B A, 吕宪国, 等. 2013. 农田开垦对三江平原湿地土壤种子库影响及湿地恢复潜力[J]. 生态学报, 33(1):205-213.
闫瑞瑞, 卫智军, 辛晓平, 等. 2011. 放牧制度对荒漠草原可萌发土壤种子库的影响[J]. 中国沙漠, 31(3): 703-708.
于洁, 高丽, 闫士坚, 等. 2015. 库布齐沙漠东段不同演替阶段沙丘土壤种子库变化特征[J]. 中国草地学报, 37(4): 80-85.
张涛, 田长彦, 孙羽, 等. 2006. 古尔班通古特沙漠地区短命植物土壤种子库研究[J]. 干旱区地理, 29(5): 675-681.
赵凌平, 程积民, 苏纪帅. 2012. 土壤种子库在黄土高原本氏针茅草地群落长期封禁演替过程中的作用[J]. 草业学报, 21(3): 38-44.
赵学勇, 张春民, 左小安, 等. 2009. 科尔沁沙地沙漠化土地恢复面临的挑战[J]. 应用生态学报, 20(7): 1559-1564.
猜你喜欢
数学物理学报(2022年5期)2022-10-09
昆明医科大学学报(2022年2期)2022-03-29
军事文摘(2021年18期)2021-12-02
食品安全导刊(2021年20期)2021-08-30
内蒙古林业(2021年6期)2021-06-26
幼儿智力世界(2021年4期)2021-05-17
河北画报(2020年8期)2020-10-27
河南科学(2020年3期)2020-06-02
科普童话·百科探秘(2018年7期)2018-09-13
浙江大学学报(工学版)(2016年2期)2016-06-05
