
青海超净区高寒草甸土壤有机碳及养分分布特征
2018-05-11 08:04:28王金贵李希来李宗仁刘育红盛海彦
生态环境学报 2018年2期
王金贵 ,李希来 ,李宗仁,刘育红,盛海彦*
1.青海大学农牧学院,青海 西宁 810016;2.省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016
土壤碳库是陆地生态系统中贮量最大的碳库(Smith,2010),据研究报道,全球土壤中的总碳量约为14×1017~15×1017g,这一贮量是陆地生物总碳量(5.6×1017g)的 2.5~3倍(刘淑丽等,2016)。每年约占大气中CO2贮量10%的碳是通过土壤排放到大气的(Raich et al.,2002),每年土壤碳排放量约为全球化石燃料燃烧碳排放量的11倍(Marland et al.,2000)。氮不仅是大气圈中含量最丰富的元素,而且也是各种植物生长和发育所需的大量营养元素之一;同时也是影响陆地生态系统初级生产力、结构和功能的主要限制性营养元素之一(Burke et al.,1997),在草原生态系统乃至全球碳氮循环中起着至关重要的作用(李明峰等,2005)。在草原生态系统中,土壤全氮不仅是主要的土壤肥力指标,也是土壤氮素肥力的基础;土壤全氮含量受植被状况、环境条件和草地利用等因素的影响(薛晓娟等,2009)。
磷是植物有机合成必不可少的重要营养元素之一(宋雄儒等,2015),是草原生态系统生产力的主要限制因素(Lovelock et al.,2007)。与林地、农田等其他生态系统一样,在草原生态系统中,土壤全磷和速效磷含量被作为重要的土壤肥力指标(宋雄儒等,2015),其分布和储量对草原生态系统功能的正常发挥具有重要作用(Jobbágy et al.,2000)。钾素不仅在土壤中含量最高,而且也是植物体吸收最多的必需营养元素之一(陈淑燕等,2013)。钾具有促进植物生理代谢和增强抗性的功能,同时还具有促进植物吸收和利用氮素的作用。虽然土壤中的全钾含量远高于土壤中的氮、磷含量,但并不代表土壤的供钾能力强,因为土壤中钾离子绝大多数以难溶状态存在,所以即使土壤中钾贮量很高,草地仍有可能供钾不足(鲍士旦,2007)。因此,不仅要了解土壤全钾含量,还有必要了解土壤速效钾含量的丰缺状况,以便指导草地恢复及生产实践。
青海省河南蒙古族自治县被联合国科教文组织誉为亚洲四大超净区之一,被国家农业部确定为全国最大有机畜产品生产基地,被国家环保部确定为国家级有机乳品生产基地。其“超净区”的含义有三:一是该区域没有工业生产所产生的污水、废气及残渣等污染;二是该区域没有农业生产所产生的面源污染;三是该区域绝对禁止使用农药化肥。然而,长期不用化肥,且牲畜粪便一般作为牧民的燃料资源被捡走,并没有归还到土壤中,该地草地土壤中的氮磷钾等养分除了大气沉降外,只有输出没有人为输入,这一生产管理方式是否会影响到该地区草原生产的可持续性?鉴于此,本研究分析了青海省河南县草甸土壤有机碳、全氮、全磷和全钾及速效养分含量的变化特征,为制定合理的草原管理制度、维持草原可持续发展和健康评价提供科学依据。
1 材料与方法
1.1 研究区自然概况
青海省河南蒙古族自治县气候为高原大陆性气候,属高原亚寒带湿润气候区。由于海拔较高,地势复杂,受季风影响强烈,所以呈现出比较明显的高原大陆性气候特点。每年 5—10月温暖、多雨,11月—次年4月寒冷、干燥、多大风天气。春秋时日短,四季不分明,无绝对无霜期。年均气温为11.9 ℃,年降水量为597.1~615.5 mm,年平均蒸发量为1349.7 mm。常年风向为西北风,最大风速达到23.7 m·s,年平均风速为2.6 m·s。年均积雪为55.3 d,最大积雪厚度为31 cm。日照率为57.58%~58.15%,略低于省内西部地区。植被主要包括禾本科、莎草科植物及多种其他科属双子叶植物。
1.2 研究方法
1.2.1 土样样品采集
采样点选取具有代表性且地上植被完整的草地。样地分布在青海省黄南藏族自治州河南蒙古族自治县的多松乡、塞尔龙乡、宁木特镇、柯生乡、优干宁镇 5个乡镇。分别采集矮嵩草(Kobresia humilis)型草甸、灌丛型草甸、禾草型草甸、沼泽型草甸和杂类草型草甸0~10 cm表层土壤样品各30份,共采集150份土样,每份土样由六点取样混合而成。去除砾石、植物残体和根系后,留取1 kg左右土样装于自封袋中带回实验室,经自然风干后,磨细、过筛、装瓶备用。高寒草甸不同草地型基本情况如表1所示。
1.2.2 测定方法
土壤有机碳测定采用重铬酸钾外加热法;土壤全氮测定采用半微量开氏法;全磷测定采用钼锑抗比色法;全钾测定采用氢氧化钠熔融-火焰光度法测定;土壤碱解氮采用碱解扩散法测定;速效磷采用碳酸氢钠浸提-钼锑抗比色法,分光光度计测定;速效钾采用1 mol·L-1乙酸铵浸提-火焰光度法测定(鲍士旦,2007)。
1.2.3 数据分析
运用SPSS 16.0软件对不同草甸土壤有机碳、全氮、全磷、全钾、速效氮、速效磷和速效钾数据采用随机模型进行方差分析,并采用新复极差法进行处理间的多重比较。
2 结果与分析
2.1 不同草地类型表层土壤有机碳含量的分布
由表2可知,高寒草甸土壤中有机碳平均含量的高低顺序为灌丛型草甸>沼泽型草甸>杂类草型草甸>矮嵩草型草甸>禾草型草甸,但5种不同类型草地有机碳含量最大值出现在杂类草型草甸,为184.91 g·kg-1;最小值出现在矮嵩草型草甸,为18.10 g·kg-1。不同类型草甸土壤有机碳变异系数大小顺序为沼泽型草甸>杂类草型草甸>矮嵩草型草甸>灌丛型草甸>禾草型草甸。根据变异系数的大小可粗略估计变量的变异程度:弱变异性,Cv<10%;中等变异性,Cv=10%~100%;强变异性,Cv>100%(王绍强等,2001)。由表 2可知,不同草甸土壤有机碳含量变异系数分布范围在26.28%~53.17%之间,属于中等变异程度。通过方差分析可知,在 5%水平上有机碳含量在灌丛型草甸、杂类草型草甸和沼泽型草甸3种类型草甸中差异不显著,但矮嵩草型草甸和禾草型草甸在5%水平上达到显著。

表1 不同类型草甸基本情况Table1 Conditions of different types of meadow

表2 不同类型草甸土壤有机碳含量Table2 Soil organic carbon content ofdifferent types ofmeadow
土壤中有机碳的输入主要取决于植物残体的形成和凋落分解,输出主要取决于土壤有机碳的分解和淋溶,所以土壤中有机碳输入与输出过程的强弱决定着土壤中有机碳含量的高低(Scholes et al.,1997)。不同的植被类型和植物功能类型均会影响土壤有机碳含量(Jackson et al.,2000)。本研究表明,灌丛型草甸、沼泽型草甸和杂类草型草甸土壤中含有较高的有机碳,而矮嵩草型草甸和禾草型草甸土壤有机碳含量相对较低,这一结果与艾丽等(2010)的研究结果基本一致。不同类型草甸地上生物量高低顺序为沼泽型草甸>灌丛型草甸>杂类草型草甸>矮嵩草型草甸>禾草型草甸,与土壤有机碳含量顺序基本一致。高寒灌丛型草甸和沼泽型草甸植被生产力较高,每年归还到土壤中的植物残体较多。另外,河南县11月—次年4月有近半年时间都处于寒冷、干燥和多风天气,所以植物残体分解相对较慢。这就导致了有机质输入过程强于输出过程,因此土壤有机碳含量相对较高。此外,灌丛型草甸中灌木残体中的木质素含量比草本植物残体中的高,所以灌丛型草甸土壤中有机质分解较慢,较稳定(傅华等,2005)。总之,在草原生态系统中,土壤有机碳含量主要受气候要素、植被类型(刘楠等,2010)、海拔和土壤特性以及人类活动的综合影响(顾振宽等,2012),从而导致土壤有机碳累积、分解和淋溶输出程度的不同(王常慧等,2004)。
2.2 不同草地类型表层土壤全氮含量分布
由表3可知,不同类型草甸土壤全氮含量高低顺序为灌丛型草甸>杂类草型草甸>沼泽型草甸>矮嵩草型草甸>禾草型草甸,全氮含量大小变化顺序与有机碳排序比较相似,其中只有杂类草型草甸和沼泽型草甸不同。矮嵩草型草甸全氮含量最小值和最大值非常接近,其变异系数也最小,为 7.93,属于弱变异程度。其他 4种类型草甸土壤全氮含量的变异系数为20.01~46.33,均属于中等变异程度。5种类型草甸土壤全氮含量差异不显著(F=1.512,P=0.271),其中灌丛型草甸土壤全氮含量最大,为7.14 g·kg-1;禾草型草甸土壤全氮含量最小,为5.52 g·kg-1。
与土壤有机碳一样,土壤全氮含量不仅受到温度和降水量等环境因子的影响,也同样与土壤特性、植被特征、海拔、土地利用方式及人类的干扰程度等因素有关(王长庭等,2005)。Smith et al.(2002)对美国华盛顿东南部半干旱区500 m海拔梯度范围内土壤氮含量的研究表明,草地植被下土壤氮含量比其他植被高;Jackson et al.(2000)对半干旱区不同植被类型土壤中氮含量的研究表明,灌木植被土壤中氮分布比草地植被深。本研究表明,灌丛型草甸、杂类型草甸和沼泽型草甸中全氮含量较高,矮嵩草型草甸和禾草型草甸中相对较低。这可能是因为,灌丛型草甸中地表残体输入量较多,有利于土壤有机质的积累。5种不同草地类型土壤中全氮含量的分布与上述土壤有机碳含量分布相似。高洋等(2015)对青藏高原高寒草甸土壤特性的研究表明,土壤氮素含量的变化与土壤有机质含量变化相似,与本研究结果一致。沼泽型草甸生产力高,虽然输人量较多,但土壤水分过多且温度较低,致使微生物活性较弱,减缓了对氮的利用,有利于氮的积累。矮嵩草型草甸和禾草型草甸地上输入量相对较少,故有机质积累也相对较少。另外,不同类型草地土壤中根系和地表调落物中碳氮比和微生物区系不同致使土壤氮分解不同也是重要原因之一(艾丽等,2010)。

表3 不同类型草甸土壤全氮含量Table3 Soil total nitrogen content ofdifferent types ofmeadow
2.3 不同类型草甸表层土壤全磷含量分布
由表4可知,不同类型草甸土壤全磷含量由低顺序为杂类草型草甸>灌丛型草甸>矮嵩草型草甸>禾草型草甸>沼泽型草甸,其平均含量近似,差异不显著(F=0.016,P=0.999),其中,杂类草型草甸土壤全磷含量最大,为 2.00 g·kg-1;沼泽型草甸土壤全磷含量最小,为 1.93 g·kg-1。在所有样本中土壤全磷含量最大值和最小值均出现在杂类草型草甸,分别为 2.86 g·kg-1和 0.93 g·kg-1。5 种草甸型土壤全磷含量的变异系数为17.01~26.64,其变异程度均属于中等变异性。
天然草原土壤中的磷主要来自于成土母质,因为天然草原生态系统与农田生态系统不同,土壤中的磷除来自大气沉降外,没有人为方式的输入(宋雄儒等,2015);其含量主要受土壤母质和成土作用的影响(李耀等,2010)。另外,研究表明,海拔、土地利用方式及坡度等因素也可以通过影响气象因素或土壤侵蚀程度而间接影响土壤磷含量(欧勇胜等,2005)。本研究结果表明,杂类草型草甸和灌丛型草甸土壤中磷含量比其他 3种草甸土壤高,但含量差异不显著,究其原因可能与放牧强度有关。杂类草型草甸和灌丛型草甸采食压力一般比矮嵩草草甸、禾草型草甸和沼泽型草甸小,即放牧强度较小,所以杂类草型草甸和灌丛型草甸土壤全磷含量相对较高。由此推断,禁牧或适度放牧可以有效保持土壤磷含量,有利于土壤养分的累积(李耀等,2010)。
2.4 不同类型草甸表层土壤全钾含量分布
由表5可知,随着草地类型的变化,不同类型草甸土壤全钾含量高低顺序为矮嵩草型草甸>禾草型草甸>杂类草型草甸>灌丛型草甸>沼泽型草甸,差异不显著(F=0.289,P=0.878)。矮嵩草型草甸和禾草型草甸土壤全钾含量的变异系数分别为 7.33和5.76,属于弱变异程度;其余灌丛型草甸、杂类草型草甸和沼泽型草甸土壤全钾含量的变异系数分别为10.74、10.75和13.15,均属于中等变异性。
不同类型草甸土壤中全钾含量远高于全氮和全磷,但98%以上的土壤全钾均以无机形态存在于土壤中。研究表明,土壤全钾对放牧干扰的反应比土壤全磷更为敏感,过度放牧会导致土壤理化性质退化,从而造成土壤颗粒变粗,而土粒越粗,土壤胶体性状越弱,土壤对钾等离子的吸附性能就越差,从而容易引起土壤全钾的流失,导致草地过牧区的土壤钾含量较低(彭景涛,2012)。河南县 5种类型草甸土壤全钾含量较高,但各类型草甸间差异不显著,说明河南县草地放牧强度适宜,并未导致土壤理化性质退化。

表4 不同类型草甸土壤全磷含量Table4 Soil total phosphorous content ofdifferent types ofmeadow
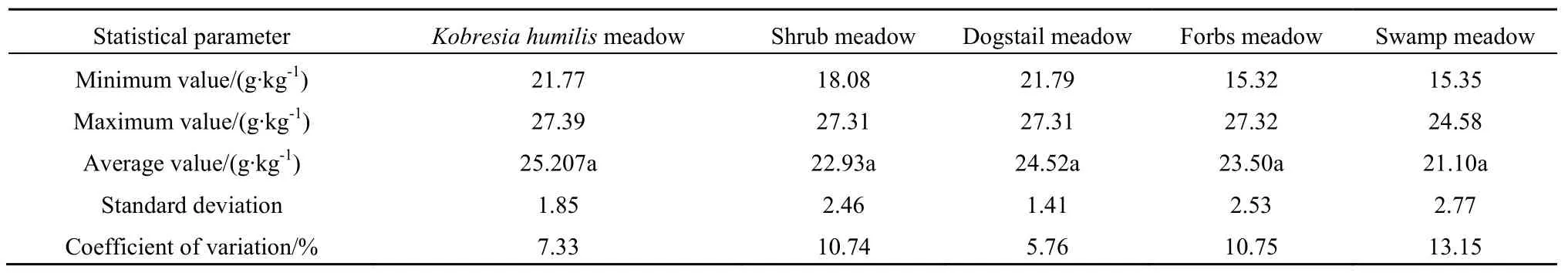
表5 不同类型草甸土壤全钾含量Table5 Soil total potassium content ofdifferent types ofmeadow
2.5 不同草地类型表层土壤速效养分含量分布
由表6可知,灌丛型草甸土壤碱解氮含量最大,为 438.72 mg·kg-1;禾草型草甸最低,为 391.10 mg·kg-1,其高低顺序与土壤全氮一致。灌丛型草甸、杂类草型草甸和沼泽型草甸3种草甸土壤碱解氮含量在 5%水平上差异不显著,矮嵩草型草甸、禾草型草甸和沼泽型草甸土壤碱解氮含量差异在 5%水平上也不显著;矮嵩草型草甸和禾草型草甸与灌丛型草甸和杂类草型草甸之间土壤碱解氮含量差异在 5%的水平上显著。不同类型草甸土壤速效磷含量高低顺序为沼泽型草甸>杂类型草草甸>禾草型草甸>灌丛型草甸>矮嵩草型草甸,且5种类型草甸土壤速效磷含量差异均不显著(F=1.274,P=0.352);其中,沼泽型草甸土壤速效磷含量最大,为 13.79 mg·kg-1;矮嵩草型草甸土壤速效磷含量最小,为10.32 mg·kg-1。不同类型草甸土壤速效钾含量以杂类草型草甸最大,为350.94 mg·kg-1;沼泽型草甸最小,为246.25 mg·kg-1。矮嵩草型草甸、禾草型草甸和杂类草型草甸土壤全钾含量差异在 5%水平上不显著,灌丛型草甸和沼泽型草甸土壤全钾含量差异在 5%水平上也不显著;矮嵩草型草甸、禾草型草甸和杂类草型草甸与灌丛型草甸和沼泽型草甸土壤全钾含量在5%水平上差异显著。

表6 不同类型草甸土壤速效养分含量Table6 Soil available nutrients content ofdifferent types ofmeadow g·kg-1
由于目前有关研究草地土壤速效养分丰缺指标的文献较少,本研究参考农田土壤碱解氮和速效磷丰缺等级指标进行分析。供氮水平:极低(N<60 mg·kg-1)、低(60~100 mg·kg-1)、中(100~125 mg·kg-1)、高(125~160 mg·kg-1)和极高(N>160 mg·kg-1)。供磷水平:极低(P<11 mg·kg-1)、低(11~20 mg·kg-1)、中(20~26 mg·kg-1)、高(26~34 mg·kg-1)和极高(P>34 mg·kg-1。刘辰琛等,2010)。根据上述标准,河南县5种类型草甸土壤供氮潜力极高,但供磷潜力较低。通常用1 mol·L-1乙酸铵浸提液浸提测定的速效钾含量作为反映土壤供钾能力和钾素水平的主要评价指标(谢建昌,2000):供钾水平极低(K<30 mg·kg-1)、中等(30~60 mg·kg-1)、高(100~160 mg·kg-1)和极高(K>160 mg·kg-1。章明清等,2008)。按照上述的评价标准,综合中国其他地区土壤速效钾的丰缺指标,初步认为,河南县的草地土壤供钾潜力极高,已达到富钾标准。
2.6 不同类型草甸土壤有机碳与全氮和碱解氮含量的关系
土壤有机碳是土壤质量构成的主要因素(杨合龙等,2015),也是土壤健康质量的关键指示指标(Percival et al.,2000),同时在全球生态系统碳循环过程中起着极其重要的作用(Joseph et al.,2011)。草地土壤中的全氮主要来源于有机质中的氮素,且主要以有机态氮的形态存在于土壤中,占全氮含量的 82%以上,土壤中全氮与有机碳含量的比值为1∶10~1∶12(陈开华等,2009)。本研究对土壤有机碳含量与土壤全氮含量和土壤碱解氮含量之间的相关关系进行了分析,结果表明,土壤有机碳与土壤全氮之间存在极显著的正相关关系,其回归方程为y=0.0729x+1.2593,相关系数r=0.9725(图 1);土壤有机碳与土壤碱解氮之间同样存在极显著正相关关系,其回归方程为y=3.1101x+194.38,相关系数r=0.7997(图2)。从相关性分析结果可知,土壤全氮和碱解氮含量的变化特征与土壤有机碳含量的变化特征相一致,这一结果与曹丽花等(2011)和高洋等(2015)的研究结果相一致。
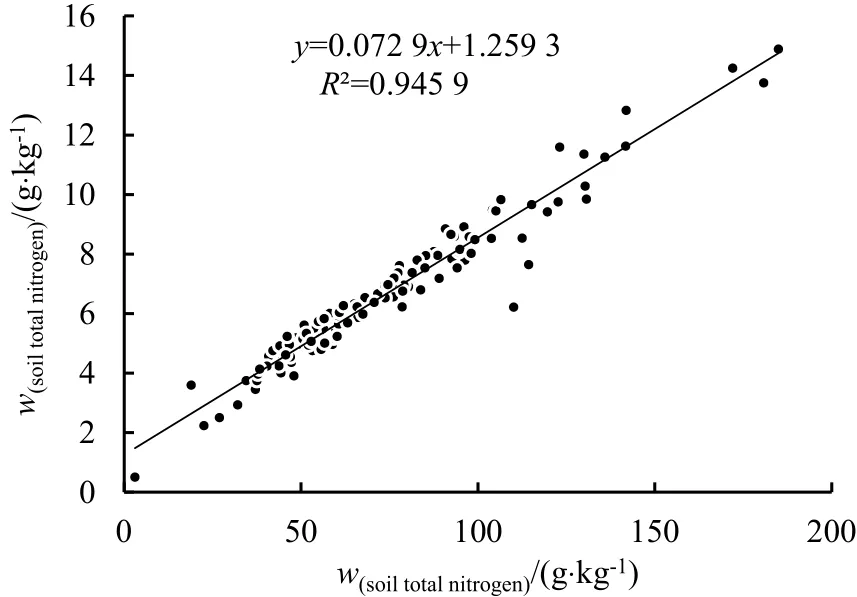
图1 土壤有机碳与全氮的关系图Fig.1 Relationship between soil organic carbon and total nitrogen

2 土壤有机碳与碱解氮的关系Fig.2 Relationship between soil organic carbon and available nitrogen
草原生态系统中,有效性氮素是限制初级生产力的首要资源(Vitousek et al.,1991),也是决定生态系统物种组成的主要因子(Tilman,1988)。相关研究表明,退化草甸生物量随着氮肥施用量的增加呈显著增加趋势(黄军等,2009)。由图1、图2可知,土壤有机碳与土壤全氮和土壤碱解氮均呈极显著相关;土壤有机碳含量越高,土壤全氮和碱解氮含量就越高。颜淑云等(2010)研究也表明,高寒草地土壤全氮和碱解氮与土壤有机碳之间呈极显著正相关关系。这表明土壤有机碳与土壤全氮和碱解氮之间存在着密切的关系,退化草地中施有机肥有利于补充速效氮含量,促使植物生长。
3 结论
(1)河南县5种不同类型草甸土壤有机碳平均含量以灌丛型草甸最高,禾草型草甸最低,有机碳含量变化属于中等变异程度。5种不同类型草甸土壤全氮含量差异不显著,其中灌丛型草甸土壤全氮含量最大,禾草型草甸最小。土壤全磷含量以杂类草型草甸最大,沼泽型草甸最小。土壤全钾含量以矮嵩草型草甸最大,沼泽型草甸最小。
(2)根据土壤速效养分丰缺标准,河南县5种不同类型草甸土壤供氮供钾潜力极高,已达到富钾标准,但供磷潜力较低,草地外源养分补充策略以施磷为主。
(3)5种不同类型草甸土壤有机碳与土壤全氮和土壤碱解氮均呈极显著正相关关系,其回归方程分别为:y=0.0729x+1.2593,相关系数r=0.9725;y=3.1101x+194.38,相关系数r=0.7997。
参考文献:
BURKE I C, LAUENROTH W K, PATTON W J. 1997. Regional and temporal variation in net primary production and nitrogen mineralization in grasslands [J]. Ecology, 78(5): 1330-1340.
JACKSON R B, SCHENK H J, JOBBÁGYE G, et al. 2000. Belowground consequences of vegetation change and their treatment in models [J].Ecological Application, 10(2): 470-483.
JOBBÁGY E G, JACKSON R B. 2000. The vertical distribution of soil organic carbon and its relation to climate and vegetation [J]. Ecological Applications, 10(2): 423-436.
JOSEPH M C, THEODORE M G. 2011. Soil moisture controls on temperature sensitivity of soil organic decomposition for a mesic grassland [J]. Soil Biology and Biochemistry, 43(2): 455-457.
LOVELOCK C E, FELLER I C, BALL M C, et al. 2007. Testing the growth rate vs. geochemical hypothesis for latitudinal variation in plant nutrients [J]. Ecology Letters, 10(12): 1154-1163.
MARLAND G,BODEN T A,ANDRES R J.2000.Global regional and national CO2emissions[C].In trends:A compendium of date on global change.Carbon dioxide information analysis center,oak ridge national laboratory,US department of energy,Oak Ridge,Tennessee.
PERCIVAL H J, PARFITT R L, SCOTT N A. 2000. Factors controllingsoil carbon level in New Zealand grassland [J]. Soil Science Society of America Journal, 64: 1623-1630.
RAICH J W, POTTER C S, BHAGAWATI D. 2002. Interannul variability in global soil respiration [J]. Global Change Biology, 8: :800-812.
SCHOLES M C, POWLSON D, TIAN G. 1997. Input control of organic matter dynamics [J]. Geoderma, 79(1-4): 25-47.
SMITH J, HALVORSON J J, BOLTON J H. 2002. Soil properties and microbial activity across a 500m elevation gradient in a semi-arid environment [J]. Soil Biology & Biochemistry, 34(11): 1749-1757.
SMITH P, FANG C. 2010. Carbon cycle: A warm response by soils [J].Nature, 464(7288): 499-500.
TILMAN D. 1988. Plant Strategies and Dynamics and Structure of Plant Communities [M]. Princeton: Princeton UniversityPress: 26-28.
VITOUSEK P M, HOWARTH R W. 1991. Nitrogen limitation on land and in the sea: How can it occur? [J]. Biogeochemistry, 13(2): 87-115.
艾丽, 吴建国, 刘建泉, 等. 2010. 土壤有机碳和全氮含量及其与海拔、植被和气候要素的关系——以祁连山中段北坡为研究对象[J]. 中国园艺文摘, (3): 27-33.
鲍士旦. 2007. 土壤农业化学分析方法[M]. 北京: 中国农业出版社.
曹丽花, 刘合满, 赵世伟. 2011. 退化高寒草甸土壤有机碳分布特征及与土壤理化性质的关系[J]. 草业科学, 28(8): 1411-1415.
陈开华, 殷恒霞, 刘俊英, 等. 2009. 高寒草甸不同植被类型土壤全氮含量变化动态分析[J]. 生态环境学报, 18(6): 2321-2325.
陈淑燕, 张德罡. 2013. 不同退化阶段高寒草甸草地土壤钾素的变化分析[J]. 草原与草坪, 33(3): 73-77.
傅华, 裴世芳, 张洪荣. 2005. 贺兰山西坡不同海拔梯度草地土壤氮特征[J]. 草业学报, 14(6):50-56.
高洋, 王根绪, 高永恒. 2015. 长江源区高寒草地土壤有机质和氮磷含量的分布特征[J]. 草业科学, 32(10): 1548-1554.
顾振宽, 杜国祯, 朱炜歆, 等. 2012. 青藏高原东部不同草地类型土壤养分的分布规律[J]. 草业科学, 29(4): 507-512.
黄军, 王高峰, 安沙舟, 等. 2009. 施氮对退化草甸植被结构和生物量及土壤肥力的影响[J]. 草业科学, 26(3): 75-78.
李明峰, 董云社, 齐玉春, 等. 2005. 温带草原土地利用变化对土壤碳氮含量的影响[J]. 中国草地, 27(1): 1-6.
李耀, 卫智军, 刘红梅, 等. 2010. 不同放牧制度对典型草原土壤中全磷和速效磷的影响[J]. 内蒙古草业, 22(1): 5-6.
刘辰琛, 吉艳芝, 张丽娟, 等. 2010. 河北清苑县冬小麦土壤速效氮、磷、钾丰缺指标的初步研究[J]. 土壤通报, 41(4): 911-915.
刘楠, 张英俊. 2010. 放牧对典型草原土壤有机碳及全氮的影响[J]. 草业科学, 27(4): 11-14.
刘淑丽, 林丽, 张法伟, 等. 2016. 放牧季节及退化程度对高寒草甸土壤有机碳的影响[J]. 草业科学, 33(1): 11-18.
欧勇胜, 张世熔, 余琼, 等. 2005. 横断山北部生态脆弱区土壤磷素空间分布特征[J]. 生态学报, 25(10): 2776-2781.
彭景涛. 2012. 青海三江源地区退化草地土壤养分元素的时空分异特征[D]. 重庆: 西南大学.
宋雄儒, 尚振艳, 李旭东, 等. 2015. 贺兰山西坡不同海拔梯度草地土壤磷特征及其影响因素[J]. 草业科学, 32(7): 1054-1060.
王常慧, 邢雪荣, 韩兴国. 2004. 草地生态系统中土壤氮素矿化影响因素的研究进展[J]. 应用生态学报, 15(11): 2184-2188.
王绍强, 朱松丽, 周成虎. 2001. 中国土壤土层厚度的空间变异性特征[J]. 地理研究, 20(2): 161-169.
王长庭, 龙瑞军, 王启基, 等. 2005. 高寒草甸不同海拔梯度土壤有机质氮磷的分布和生产力变化及其与环境因子的关系[J]. 草业学报,14(4): 15-20.
谢建昌. 2000. 钾与中国农业[M]. 南京: 河海大学出版社.
薛晓娟, 李英年, 杜明远, 等. 2009. 祁连山东段南麓不同海拔土壤有机质及全氮的分布状况[J]. 冰川冻土, 31(4): 642-649.
颜淑云, 周志宇, 秦彧, 等. 2010. 玛曲高寒草地不同利用方式下土壤氮素含量特征[J]. 草业学报, 19(2): 153-159.
杨合龙, 孙宗玖, 杨静, 等. 2015. 封育年限对伊犁绢蒿荒漠土壤活性有机碳及碳库管理指数的影响[J]. 草业科学, 32(12): 1945-1952.
章明清, 颜明娟, 林琼, 等. 2008. 土壤钾素解吸模型及其特征值的作物效应[J]. 土壤学报, 45(1): 120-129.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
湖北农业科学(2019年10期)2019-07-08 02:55:40
活力(2019年21期)2019-04-01 12:17:10
安徽农业科学(2018年15期)2018-05-14 08:59:46
科技资讯(2017年3期)2017-03-25 09:27:53
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代农业(2016年4期)2016-02-28 18:42:07
