
亚热带地区不同人工林配置下土壤微生物量碳及微生物墒的年际动态
2018-05-11 08:04易桂田王晓丽刘占锋蔡锡安傅声雷周丽霞
生态环境学报 2018年2期
易桂田 ,王晓丽,刘占锋,蔡锡安,傅声雷 ,周丽霞*
1.中国科学院华南植物园//中国科学院退化生态系统植被恢复与管理重点实验室//广东省应用植物学重点实验室,广东 广州 510650;2.中国科学院大学,北京 100049;3.省部共建三江源生态与高原农牧业国家重点实验室//青海大学,青海 西宁 810016;4.河南大学环境与规划学院,河南 开封 475004
土壤微生物是土壤肥力形成和持续发展的核心动力,它既参与土壤有机质分解释放养分,又与地上植被争夺土壤养分(沈仁芳等,2017),因此,土壤微生物的活动与土壤速效养分的来源密切相关。土壤微生物生物量可以反映土壤有机碳中具有活性的部分,该指标常被用来表示土壤微生物的群落状态和功能变化(刘占锋等,2007)。研究证实,土壤微生物生物量与土壤肥力和土壤健康的关系十分紧密(俞慎等,1999;易泽夫等,2006;Zhou et al.,2007;Bolat et al.,2015)。
由于农田土壤肥力的提高有利于增产,所以在不同施肥处理以及不同耕作制度下的农业用地中关于土壤微生物量碳的研究较多(Moore et al.,2000)。但是相关研究表明,森林生态系统中的土壤微生物量碳远远高于农业用地(Kara et al.,2008),且森林生态系统中地上植被的恢复有利于土壤生态系统的恢复,因此对于森林生态系统土壤微生物量的研究逐年增多(Jia et al.,2005;Fang et al.,2014)。森林生态系统中不同土壤类型、不同林型、不同林龄的土壤微生物量变化特征,可以反映出森林土壤的质量状况与健康水平(Bauhus et al.,1998;徐华勤等,2009);而且土壤微生物对气候、植被及土壤环境的变化十分敏感,即使在相同立地的不同树种配置下也会引起土壤微生物生物量的较大差异(李胜蓝等,2014)。此外,土壤微生物量碳在长期监测下可能更容易产生显著性差异(Hargreaves et al.,2003)。一般认为,在全球尺度上土壤微生物量碳通常随着纬度的降低而减少(Xu et al.,2013),相同气候带不同林型间的差异则表现为混交林高于纯林,阔叶林高于针叶林,这主要缘于凋落物质量与数量的差异(Thoms et al.,2010;柳杨等,2017),而不同林龄的差异则取决于不同地带的土壤类型、土壤营养环境以及地上植被的共同作用(杨凯等,2009)。研究表明,土壤微生物墒(土壤微生物量碳与土壤有机碳的比值)对土壤质量的变化更为直接和敏感,可以较早地反映出土壤肥力的变化(Sparling,1992;Wander et al.,1994;Coleman,2013)。在亚热带地区,一般认为成熟林土壤微生物墒值大于19.5 mg∙g-1,表明该自然林或人工林处于碳积累阶段;小于该值则表明该自然林或人工林处于碳损失阶段(易志刚,2005)。
目前在相同立地环境下不同森林生态系统的研究多关注于土壤微生物量碳的季节动态,对其年际变化的长期跟踪研究较为少见。而研究不同人工林配置下土壤微生物量的变化及其年际动态,有利于及时了解不同物种配置对退化土壤质量恢复的影响。为此,本研究选取广东鹤山丘陵荒坡上不同植被配置的5种人工林样地,并以自然恢复的草坡为对照,监测分析了不同配置人工林在种植第1年、第6年和第11年的土壤微生物量碳及土壤理化性质的变化,以探讨不同人工林物种配置下土壤微生物量碳的年际变化特征及其与土壤肥力的关系,为人工林的生态恢复与管理提供科学的基础数据与理论依据。
1 材料与方法
1.1 研究区域概况
研究样地选择在广东鹤山森林生态系统国家野外科学观测研究站共和样地(鹤山站,22°34'N,112°50'E)。该样地位于广东省中部,海拔10~60 m,为低丘地势,山脊平圆,坡面平缓;属南亚热带季风气候,夏季湿热,冬季干冷,干湿季交替现象明显,干季一般从10月—次年2月,湿季一般从3—9月;年均气温22.6 ℃,年均降雨量1700 mm,年蒸发量1600 mm,年辐射量4350.5 MJ∙m-2;地带性土壤为赤红壤,土壤质地以粘壤土类和壤质粘粒为主。该地区历史上为森林地带,顶极群落是亚热带季风常绿阔叶林,但由于不断增长的人类活动影响,造成植被退化、水土流失、土壤瘠瘦。为了科学地研究植被恢复和开展长期监测,2005年在广东鹤山共和镇的丘陵荒坡上建立了森林生态系统野外控制实验研究平台,构建了不同物种配置的人工林样地(图1)。共和样地建立前的丘陵荒坡经过了烧山处理,所以不同林型的土壤条件在种植初期基本一致,本研究选取了其中6种不同配置的实验样地。
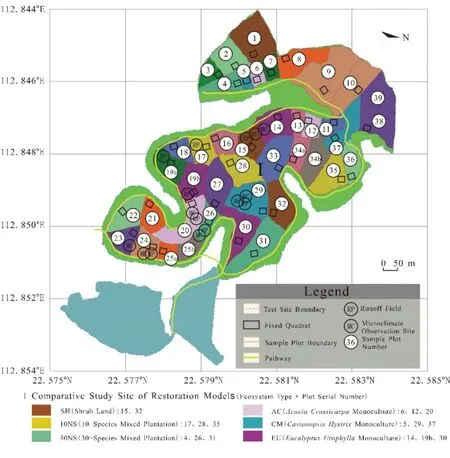
图1 实验样地分布Fig.1 Experimental ground distribution
1.2 研究方法
1.2.1 样品采集
以 2005年同期种植的 10种树种混交林(10NS)、30种树种混交林(30NS)、厚荚相思纯林(Acacia crassicarpa,AC)、红椎纯林(Castanopsis hystrix,CM)、尾叶桉纯林(Eucalyptus urophylla,EU)为研究对象,同时以自然恢复的草坡(SH)为对照样地,分别于2006年、2011年和2016年(种植第1年、第6年和第11年)的10—11月(干季)采集土壤样品进行分析。上述每种林型有3个重复样地,每个样地面积约1 hm2。为了去除土壤空间异质性,在每个样地固定样方的下坡位、中坡位和上坡位各随机选取3~5个点,去除地表凋落物和腐质层后,用内径为3 cm的不锈钢土钻,采集0~10 cm深度的土壤样品。每种林型分别采集9个土壤混合样,其中草坡只有2个重复样地,故只采集6个混合样。
1.2.2 分析方法
土样除去其中的细根、砾石等杂质,过2 mm筛后分为2份,一份带回实验室风干后进行土壤理化性质分析,一份于-4 ℃下保存用于土壤微生物量碳的测定(一周内测定完成)。土壤微生物量碳采用氯仿熏蒸法测定(Vance et al.,1987)。土壤含水量采用烘干法测定;pH值采用pH计测定,土水比为1∶2.5;土壤全氮含量采用半微量开氏法测定;土壤有机碳含量采用重铬酸钾氧化-外加热法测定(孙鸿烈等,1996)。
1.3 数据分析
数据经过Excel 2013整理后,用R3.4.0及SPSS 20.0进行统计分析。柱状图用SigmaPlot 12.3绘制。箱线图法剔除异常值,此过程运用了boxplot.stats ()函数;不同林龄与不同林型之间环境变量的差异性用单因素方差分析及Tukey组间检验分析,此过程运用了multcomp包中的glht ()函数;土壤微生物量碳及土壤微生物墒与环境变量的关系采用Spearman相关性分析并运用psych包中的corr.test()函数进行显著性检验分析,双因素分析于 SPSS 20.0中进行。各林型每样方的土壤微生物量碳、微生物墒及各种环境变量采用 3个样地的平均值表示。采用一般线性模型和广义线性模型分析林型、林龄及其交互作用对土壤微生物量碳、微生物墒的影响。显著性水平设定为0.05。
2 结果分析
2.1 不同林型土壤理化性质的年际变化
在人工林种植第1年、6年及11年,土壤理化性质在不同林型间均无显著差异。在不同林龄间,EU及 SH土壤有机碳随林龄增加而变化的趋势不显著,而 10NS(F=51.95,P=0.0002)、30NS(F=9.634,P=0.013)、AC(F=25.66,P=0.001)以及 CM(F=8.963,P=0.016)土壤有机碳随林龄增加呈先增加后减少的趋势且差异显著,但6种林型土壤有机碳含量均在 6 a林龄时达到最高。10NS(F=26.55,P=0.001)、30NS(F=5.923,P=0.038)、AC(F=11.49,P=0.009)、CM(F=14.99,P=0.005)及SH(F=10.95,P=0.042)的可溶性有机碳均随林龄增加呈先增加后减少的趋势且表现显著性差异。土壤全氮在6种林型中也以6年生林龄含量最高,但只有AC表现了显著性差异(F=18.25,P=0.003)。土壤碳氮比在不同林龄间只有EU表现出显著差异(F=8.548,P=0.018)。6种林型的pH均呈现随着林龄的增加而逐渐上升的趋势,但只有SH(F=18.03,P=0.021)、10NS(F=13.22,P=0.006)及 30NS(F=12.71,P=0.007)表现了显著性差异(表1)。
2.2 不同林型土壤微生物量碳的年际动态
双因素分析结果显示,林型(df=5,F=3.039,P=0.023;df=2,F=4.142,P=0.023)及林龄(df=5,F=3.813,P=0.032;df=2,F=3.372,P=0.044)对土壤微生物量碳有显著影响,而林型与林龄的交互作用无显著影响。单因素方差分析结果如图2所示,在1 a和6 a林龄时,6种林型间的土壤微生物量碳均无显著差异,但到了11 a林龄时6种林型间的土壤微生物量碳产生了显著差异(df=5,F=6.543,P=0.004),表现为 10NS(220.043 mg∙kg-1)与 30NS(212.155 mg∙kg-1)两种混交林的土壤微生物量碳含量最高,AC(178.049 mg∙kg-1)与 CM(104.660 mg∙kg-1)两种纯林次之,EU与 SH土壤微生物量碳含量最低,分别为 56.912 mg∙kg-1和 64.042 mg∙kg-1;在1 a和6 a林龄时,混交林、纯林与对照草坡之间土壤微生物量碳均无显著差异,但到了11 a林龄时混交林、纯林与对照草坡之间土壤微生物量碳产生了显著差异(df=2,F=7.717,P=0.006),表现为混交林的土壤微生物量碳高于纯林和对照草坡。10NS、30NS、AC、CM及 SH土壤微生物量碳随着林龄增加呈先增加后下降的趋势但无显著差异,EU土壤微生物量碳随着林龄增加呈一直下降的趋势但未表现出显著差异,6 a林龄时土壤微生物量碳在AC中达到最高值,为266.078 mg∙kg-1(图2)。
2.3 不同林型土壤微生物墒的年际动态
双因素分析结果显示,林型(df=5,χ2=16.27,P=0.006;df=2,χ2=6.298,P=0.043)及林龄(df=5,χ2=10.174,P=0.006;df=2,χ2=6.092,P=0.048)对土壤微生物墒有显著影响,林型与林龄的交互作用无显著影响。单因素方差分析结果如图3所示,在11 a林龄时 6种林型间的土壤微生物墒值表现出显著差异(df=5,F=5.483,P=0.009),其中30NS土壤微生物墒值最高,为 11.017 mg∙g-1,其次是10NS,为 10.522 mg∙g-1,最低是 EU,为 2.811 mg∙g-1;11 a林龄时的混交林、纯林及草坡之间的土壤微生物墒值表现出显著差异性(df=2,F=5.299,P=0.019),表现为混交林最高,对照SH与纯林次之。6种林型的土壤微生物墒在林龄间均无显著性差异(图3)。
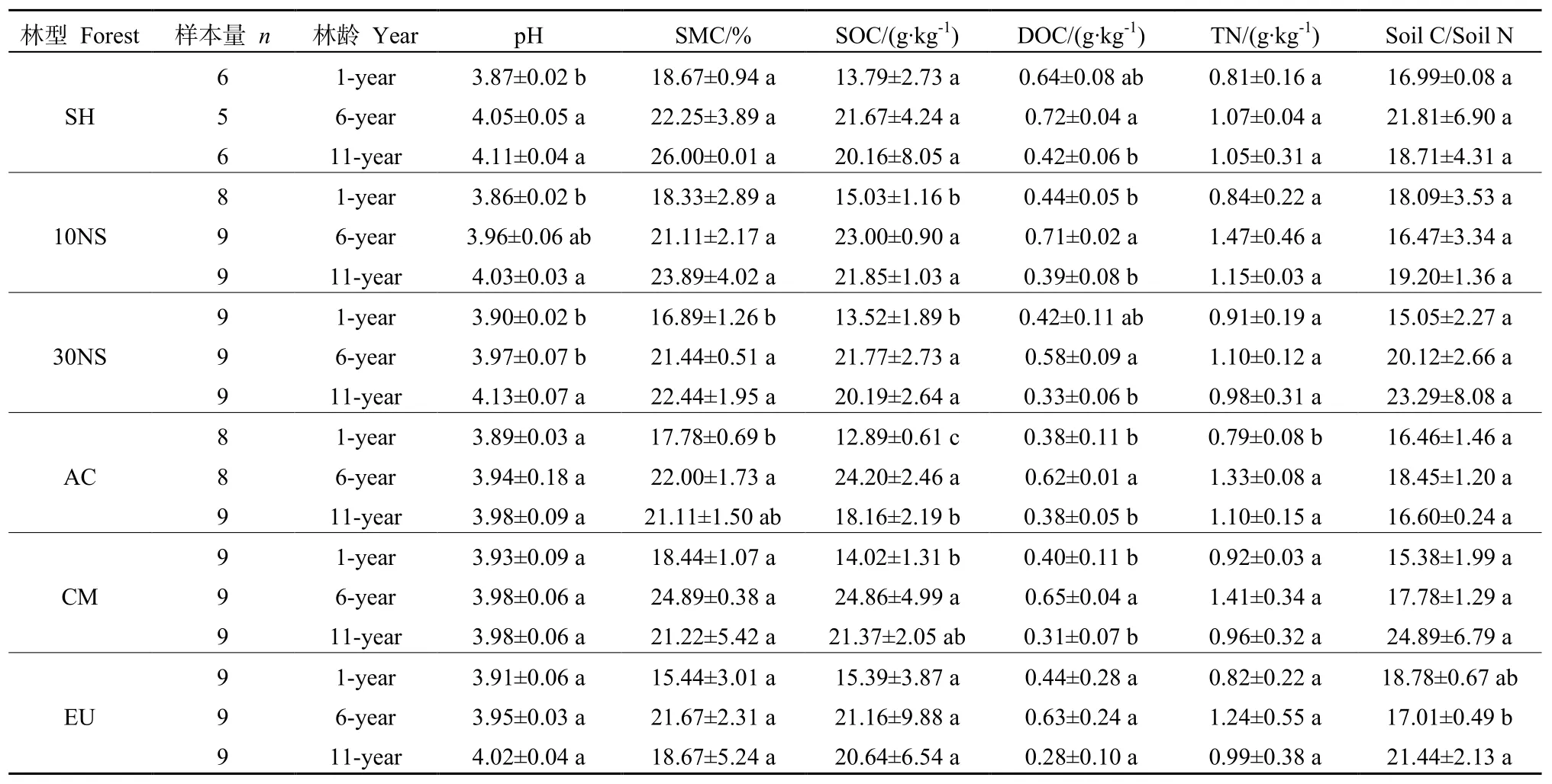
表1 不同林型土壤理化性质的年际动态Table1 Interannual dynamics of soil properties in different forest types
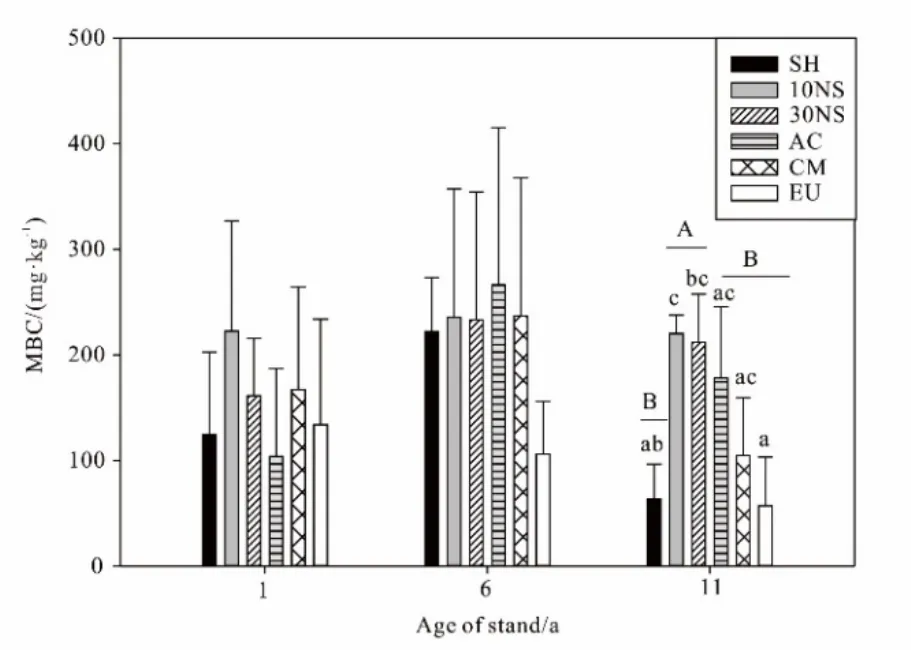
图2 不同林型土壤微生物量碳的年际动态Fig.2 Interannual dynamics of Soil microbial biomass C in different forest types

图3 不同林型土壤微生物墒的年际动态Fig.3 Interannual dynamics of Soil microbial biomass C/soil organic C in different forest types
2.4 土壤微生物碳及土壤微生物墒与土壤理化性质的相关性
研究结果表明,不同人工林土壤有机碳、可溶性有机碳及全氮的变化趋势与不同人工林(除了 EU)土壤微生物量碳变化趋势一致,均在人工林种植的第6年达到最高含量。相关性分析结果表明,土壤微生物量碳与土壤有机碳(r=0.25,P<0.01)、可溶性有机碳(r=0.23,P=0.01)及全氮(r=0.23,P=0.01)呈显著正相关,而土壤微生物墒与土壤有机碳(r=0.18,P=0.03)、土壤碳氮比(r=0.18,P=0.04)呈显著负相关,与可溶性有机碳呈显著正相关(r=0.18,P=0.04)。土壤微生物量碳及土壤微生物墒与土壤pH不存在相关性(表2),说明本研究中土壤微生物量碳及土壤微生物墒与土壤养分的变化更加密切。

表2 土壤微生物量指标与土壤理化性质的相关性分析Table2 Spearman correlations coefficients between soil microbial biomass variables and soil physicochemical properties
3 讨论
3.1 不同林型土壤微生物量碳的年际变化

图4 2006年、2011年以及2016年降水及气温的月均值Fig.4 Average monthly of precipitation and temperature in 2006, 2011 and 2016
土壤微生物量碳是土壤微生物所固定的碳的量(刘爽等,2010),是占土壤全碳的比例小却是最活跃的一部分,是土壤中易于被利用的养分库及有机物分解的动力,故其在土壤肥力和生态系统评价中起着重要的作用(俞慎等,1999;王意锟等,2012)。由图 4可知,在 2006年、2011年、2016年采样期间,研究样地降水和温度差异不大(图4),同时Cao et al.(2010)在东莞桉林土壤微生物研究中的结果显示,该地区土壤微生物量碳在干湿季并无显著差异,因此本研究以2006年、2011年及2016年干季土壤微生物量碳的数据分析土壤微生物量碳的年际动态。研究结果显示,土壤微生物量碳在年际间的变化没有显著性差异,但是存在一定的变化趋势,除EU外,其他林型土壤微生物量碳随着林龄的增加均呈先增加后下降的趋势,在6 a林龄达到最高。这可能是由于人工林林下灌草对土壤微生物生物量影响很大,但是随着人工林的恢复其郁闭度增加使得林下灌草大量减少,因此在6 a林龄后出现下降趋势(Zhao et al.,2012;Zhao et al.,2013;Zhao et al.,2014;Chen et al.,2017)。EU土壤微生物量碳随着林龄增加呈下降趋势且其含量相对较低,根据Chen et al.(2017;2015)研究可知,EU是快速生长型树种,其地上生物量随着林龄增加呈上升趋势且显著高于其他林型,说明EU地上植被的生长与土壤微生物量碳的固定可能存在竞争关系。本研究显示,11 a林龄时土壤微生物量碳在不同林型间表现出显著差异,总体趋势是10NS、30NS>AC、CM>EU、SH。有研究表明,植被通过根系分泌物以及凋落物向土壤提供了碳氮元素相关养分,影响了土壤有机质的输入从而影响土壤性质(刘占锋等,2007;Rutigliano et al.,2004),且凋落物的不同搭配能显著提高土壤微生物量碳。因为植被种类越多,凋落物资源的异质性就越大,可为不同种类的微生物提供不同的营养物质,有利于土壤微生物的大量繁殖及多样性的形成(王春阳等,2011;王意锟等,2012),所以混交林的土壤微生物量显著高于纯林(Fang et al.,2016)。AC土壤微生物量碳相对较高,可能与AC作为固氮树种所营造的土壤环境更有利于土壤微生物量碳的固定有关(Huang et al.,2014)。SH及EU土壤微生物量碳在11 a林龄时明显低于其他混交林和纯林,说明地上植被的种植及地上植被类型均会间接影响土壤微生物量。研究结果反映出土壤微生物量碳在年际间的变化波动较大,这可能与人工植被种植初期林相不够稳定,植被和土壤尚未达到成熟水平有关(Insam et al.,1988)。
3.2 不同林型土壤微生物墒的年际变化
土壤微生物墒反映了土壤中活性有机碳所占比例,可作为土壤质量研究的有效指标(张帆等,2009;徐华勤等,2009;徐一兰等,2016)。Cheng et al.(2013)和刘占锋等(2007)研究表明,土壤微生物量碳与土壤有机碳的比值越高,则土壤活性有机碳的积累越多,越有利于土壤微生物的生长,反之则表明惰性有机质的积累使得供给土壤微生物的有机质减少。刘占锋等(2007)结果显示,土壤微生物墒在恢复进程中呈下降趋势,本研究结果显示,土壤微生物墒在年际间的变化差异性并不显著,由此可以推断林型对于土壤微生物墒的作用可能更大。已有研究表明,鼎湖山自然成熟林土壤微生物墒值为19.5 mg∙g-1,大于该值表示林分土壤处于碳积累阶段(易志刚等,2003;易志刚等,2005),由此推测本研究中11 a林龄的人工林尚未进入植被恢复的成熟阶段。11 a林龄后是否会呈上升的变化趋势还有待进一步监测分析。本研究中不同林型之间土壤微生物墒在11 a林龄产生差异,混交林土壤微生物墒显著高于纯林,这可能是因为地表凋落物输入的质量和数量存在差异,使得混交林土壤微生物量碳和土壤养分含量与其他纯林间产生了差异(Thoms et al.,2010),这与鼎湖山凋落物分解较快的季风阔叶林土壤微生物墒值低于凋落物分解较慢的马尾松林的结果不完全一致,也说明共和样地的人工林仍处于以碳损失为主的非成熟阶段。
3.3 土壤微生物量碳与土壤养分变化的关系
大量研究显示,土壤微生物量碳与土壤有机碳、可溶性有机碳及全氮关系密切(Li et al.,2004;黄宇等,2004;汪文霞等,2006;赵彤等,2013),本研究结果表明,随着林龄的增加,不同配置人工林(除了 EU)土壤微生物量碳与土壤有机碳、可溶性有机碳及全氮均呈先增加后减少趋势且存在显著正相关,并在6 a林龄时达到最高。然而,Jia et al.(2011)研究显示,土壤微生物量碳和土壤有机碳随林龄增加均呈增加趋势,在20 a林龄时达到最高,而全氮则在20 a林龄达到最低后在30 a时达到最高,这说明相同气候带不同研究点与植被类型,其生长所需的土壤养分在种类和数量上也存在差异。本研究中固氮树种AC的土壤微生物量碳高于其他纯林,而相关研究显示施加氮肥对土壤微生物量碳并无显著影响(Moore et al.,2000),这说明添加氮并不能直接导致土壤微生物量的增加,土壤微生物量的变化可能与植被生长过程中根系分泌物及凋落物的输入对土壤养分状况的改变有关(Wang et al.,2004)。随着林龄的增加,速生型树种EU土壤微生物量碳及土壤有机碳、可溶性有机碳及全氮均无显著变化,且土壤微生物量碳呈下降趋势,说明这 11年来桉树林对土壤有机碳、全氮的恢复作用不显著,且供给土壤微生物的碳较少。一般认为,人工林可通过施肥来改善植物和土壤微生物对土壤养分的竞争关系,但其反应与调节机制还有待进一步研究。土壤碳氮比是有机质与氮素总质量的比值,是表征土壤有机质组成和质量程度的一个重要指标(王绍强等,2008)。本研究结果显示,30NS和CM土壤碳氮比,10NS、30NS和SH土壤含水量与土壤pH随着林龄的增加呈增加趋势,说明随着植被的恢复以及土壤微生物活性的增加,这些林型的土壤养分结构逐步趋于稳定,土壤质量也逐步提高。本研究中土壤pH与土壤微生物量碳及微生物墒无显著相关性,说明在本研究样地中土壤有机碳及全氮才是决定土壤微生物量的重要因素。
4 结论
综上所述,在人工林种植初期(1、6、11 a),随林龄增加土壤微生物量碳除在尾叶桉纯林中呈下降趋势外,在其他林型中均呈先上升后下降的趋势,但年际间的变化趋势均不显著。土壤微生物量碳在11 a林龄不同林型间存在显著差异,总体趋势为混交林>红椎纯林、厚荚相思纯林>桉林、草坡,这说明植被恢复早期(1~6 a林龄),地上植被通过凋落物的输入促进了土壤微生物的活性,且不同树种的凋落物匹配能为土壤微生物提供更多的基质及机遇,混交林更有利于土壤微生物多样性的形成,因此土壤微生物量碳含量也更高;但由于土壤微生物量碳的固定与植物生长存在竞争关系,故植被生长到一定阶段(11 a林龄)后土壤微生物量碳有所下降,这可能与11 a林龄林分和土壤尚未达到成熟稳定的水平有关。
参考文献:
BAUHUS J, PARE D, COTE L. 1998. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest [J]. Soil Biology & Biochemistry, 30(8-9): 1077-1089.
BOLAT I, SENSOY H, OZER D. 2015. Short-term changes in microbial biomass and activity in soils under black locust trees (Robinia pseudoacacia L.) in the northwest of Turkey [J]. Journal of Soils and Sediments, 15(11): 2189-2198.
CAO Y S, FU S L, ZOU X M, et al. 2010. Soil microbial community composition under Eucalyptus plantations of different age in subtropical China [J]. European Journal of Soil Biology, 46(2):128-135.
CHEN Y Q, LIU Z F, RAO X Q, et al. 2015. Carbon Storage and Allocation Pattern in Plant Biomass among Different Forest Plantation Stands in Guangdong, China [J]. Forests, 6(3): 794-808.
CHEN Y Q, YU S Q, LIU S P, et al. 2017. Reforestation makes a minor contribution to soil carbon accumulation in the short term: Evidence from four subtropical plantations [J]. Forest Ecology and Management,384: 400-405.
CHENG F, PENG X, ZHAO P, et al. 2013. Soil Microbial Biomass, Basal Respiration and Enzyme Activity of Main Forest Types in the Qinling Mountains [J]. PLoS One, 8(6): 1-12.
COLEMAN D. 2013. Soil Biota, soil systems and processes [M]//Encyclopedia of Biodiversity. New York: Academic Press: 305-314.
FANG X M, WANG Q L, ZHOU W M, et al. 2014. Land use effects on soil organic carbon, microbial biomass and microbial activity in Changbai Mountains of Northeast China [J]. Chinese Geographical Science,24(3): 297-306.
FANG X M, YU D P, ZHOU W M, et al. 2016. The effects of forest type on soil microbial activity in Changbai Mountain, Northeast China [J].Annals of Forest Science, 73(2): 473-482.
HARGREAVES P R, BROOKES P C, ROSS G J S, et al. 2003. Evaluating soil microbial biomass carbon as an indicator of long-term environmental change [J]. Soil Biology & Biochemistry, 35(3):401-407.
HUANG X, LIU S, WANG H, et al. 2014. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China[J]. Soil Biology & Biochemistry, 73(6): 42-48.
INSAM H, DOMSCH K H. 1988. RELATIONSHIP BETWEEN SOIL ORGANIC-CARBON AND MICROBIAL BIOMASS ON CHRONOSEQUENCES OF RECLAMATION SITES [J]. Microbial Ecology, 15(2): 177-188.
JIA G M, CAO J, WANG C Y, et al. 2005. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin,northwest China [J]. Forest Ecology and Management, 217(1):117-125.
JIA G M, ZHANG B L, WU Z R, et al. 2011. Microbial biomass and nutrients in soil at the different ages of Citrus in Three Gorges Reservoir area [J]. International Symposium on Water Resource &Environmental Protection, 3: 1694-1696.
KARA Ö, BOLAT İ. 2008. The effect of different land uses on soil microbial biomass carbon and nitrogen in Bartin Province [J]. Turkish Journal of Agriculture & Forestry, 32(4): 281-288.
LI Q C, ALLEN H L, WOLLUM A G. 2004. Microbial biomass and bacterial functional diversity in forest soils: effects of organic matter removal, compaction, and vegetation control [J]. Soil Biology &Biochemistry, 36(4): 571-579.
MOORE J, KLOSE S, TABATABAI M. 2000. Soil microbial biomass carbon and nitrogen as affected by cropping systems [J]. Biology and Fertility of Soils, 31(3-4): 200-210.
RUTIGLIANO F A, D'ASCOLI R, DE SANTO A V. 2004. Soil microbial metabolism and nutrient status in a Mediterranean area as affected by plant cover [J]. Soil Biology & Biochemistry, 36(11): 1719-1729.
SPARLING G P. 1992. RATIO OF MICROBIAL BIOMASS CARBON TO SOIL ORGANIC-CARBON AS A SENSITIVE INDICATOR OF CHANGES IN SOIL ORGANIC-MATTER [J]. Australian Journal of Soil Research, 30(2): 195-207.
THOMS C, GATTINGER A, JACOB M, et al. 2010. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest [J]. Soil Biology & Biochemistry, 42(9): 1558-1565.
VANCE E D, BROOKES P C, JENKINSON D S. 1987. An extraction method for measuring soil microbial biomass-C [J]. Soil Biology &Biochemistry, 19(19): 703-707.
WANDER M M, TRAINA S J, STINNER B R, et al. 1994. Organic and conventional management effects on biologically-active soil organic-matter pools [J]. Soil Science Society of America Journal,58(4): 1130-1139.
WANG F E, CHEN Y X, TIAN G M, et al. 2004. Microbial biomass carbon,nitrogen and phosphorus in the soil profiles of different vegetation covers established for soil rehabilitation in a red soil region of southeastern China [J]. Nutrient Cycling in Agroecosystems, 68(2):181-189.
XU X F, THORNTON P E, POST W M. 2013. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems [J]. Global Ecology and Biogeography, 22(6): 737-749.
ZHAO J, WAN S Z, FU S L, et al. 2013. Effects of understory removal and nitrogen fertilization on soil microbial communities in Eucalyptus plantations [J]. Forest Ecology and Management, 310(1): 80-86.
ZHAO J, WAN S Z, LI Z A, et al. 2012. Dicranopteris-dominated understory as major driver of intensive forest ecosystem in humid subtropical and tropical region[J]. Soil Biology & Biochemistry, 49(5):78-87.
ZHAO J, WAN S Z, ZHANG C L, et al. 2014. Contributions of Understory and/or Overstory Vegetations to Soil Microbial PLFA and Nematode Diversities in Eucalyptus Monocultures [J]. PLoS One, 9(1): e85513.
ZHOU L, DING M. 2007. Soil microbial characteristics as bioindicators of soil health [J]. Biodiversity Science, 15(2): 162-171.
黄宇, 汪思龙, 冯宗炜, 等. 2004. 不同人工林生态系统林地土壤质量评价[J]. 应用生态学报, 15(12): 2199-2205.
李胜蓝, 方晰, 项文化, 等. 2014. 湘中丘陵区4种森林类型土壤微生物生物量碳氮含量[J]. 林业科学, 50(5): 8-16.
刘爽, 王传宽. 2010. 五种温带森林土壤微生物生物量碳氮的时空格局[J]. 生态学报, 30(12): 3135-3143.
刘占锋, 刘国华, 傅伯杰, 等. 2007. 人工油松林 (Pinus tabulaeformis)恢复过程中土壤微生物生物量 C、N的变化特征[J]. 生态学报,27(3): 1011-1018.
柳杨, 何先进, 侯恩庆. 2017. 鼎湖山森林演替和海拔梯度上的土壤微生物生物量碳氮变化[J]. 生态学杂志, 36(2): 287-294.
沈仁芳, 孙波, 施卫明, 等. 2017. 地上-地下生物协同调控与养分高效利用[J]. 中国科学院院刊, 32(6): 566-574.
孙鸿烈, 刘光崧. 1996. 中国生态系统研究网络观测与分析标准方法[M]. 北京: 中国标准出版社.
王春阳, 周建斌, 夏志敏, 等. 2011. 黄土高原区不同植物凋落物搭配对土壤微生物量碳,氮的影响[J]. 生态学报, 31(8): 2139-2147.
王绍强, 于贵瑞. 2008. 生态系统碳氮磷元素的生态化学计量学特征[J].生态学报, 28(8): 3937-3947.
汪文霞, 周建斌, 严德翼, 等. 2006. 黄土区不同类型土壤微生物量碳、氮和可溶性有机碳、氮的含量及其关系[J]. 水土保持学报, 20(6):103-106.
王意锟, 方升佐, 田野, 等. 2012. 残落物添加对农林复合系统土壤有机碳矿化和土壤微生物量的影响[J]. 生态学报, 32(22): 7239-7246.
徐华勤, 章家恩, 冯丽芳, 等. 2009. 广东省不同土地利用方式对土壤微生物量碳氮的影响[J]. 生态学报, 29(8): 4112-4118.
徐一兰, 唐海明, 肖小平, 等. 2016. 长期施肥对双季稻田土壤微生物学特性的影响[J]. 生态学报, 36(18): 5847-5855.
杨凯, 朱教君, 张金鑫, 等. 2009. 不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J]. 生态学报, 29(10): 5500-5507.
易泽夫, 荣湘民, 彭建伟, 等. 2006. 微生物变量作为土壤质量评价指标的探讨[J]. 湖南农业科学, (6): 64-65, 69.
易志刚, 蚁伟民, 周国逸, 等. 2003. 鼎湖山三种主要植被类型土壤碳释放研究[J]. 生态学报, 23(8): 1673-1678.
易志刚, 蚁伟民, 周丽霞, 等. 2005. 鼎湖山主要植被类型土壤微生物生物量研究[J]. 生态环境, 14(5): 727-729.
俞慎, 李勇, 王俊华, 等. 1999. 土壤微生物生物量作为红壤质量生物指标的探讨[J]. 土壤学报, 36(3): 413-413.
张帆, 黄凤球, 肖小平, 等. 2009. 冬季作物对稻田土壤微生物量碳、氮和微生物熵的短期影响[J]. 生态学报, 29(2): 734-739.
赵彤, 闫浩, 蒋跃利, 等. 2013. 黄土丘陵区植被类型对土壤微生物量碳氮磷的影响[J]. 生态学报, 33(18): 5615-5622.
猜你喜欢
防护林科技(2022年5期)2022-08-22
江苏农业学报(2022年1期)2022-03-16
西北林学院学报(2021年6期)2021-12-07
空间科学学报(2021年2期)2021-07-21
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
林业科学(2017年3期)2017-04-27
中国学术期刊文摘(2016年1期)2016-02-13
中国学术期刊文摘(2016年8期)2016-02-13
应用海洋学学报(2015年4期)2015-11-24
