
老年肌肉衰减综合征基因表达性别差异的生物信息学分析
2018-03-30 08:36高原张伟波陈健寿崟徐平虎力
中国康复理论与实践 2018年3期
高原,张伟波,陈健,寿崟,徐平,虎力
1.上海中医药大学针灸推拿学院,上海市201203;2.上海中医药大学中医复杂系统研究中心,上海市201203
原发性肌肉衰减综合征是一组与增龄相关的骨骼肌质量和功能逐渐下降的综合征[1]。2010年欧洲老年肌肉衰减综合征工作组推荐将肌肉质量、力量、执行力同时降低,作为老年肌肉衰减综合征临床诊断的标准[2]。之后中国台湾[3]及亚洲肌肉衰减综合征工作组[1,4]针对欧洲标准提出修改建议。引起该综合征的因素大概可归结为肌肉废用、内分泌功能改变、慢性炎症、胰岛素抵抗(insulin resistance,IR)及营养缺乏[5]。骨骼肌作为人体重要的内分泌器官,参与葡萄糖、脂质等代谢过程[6]。在糖尿病人群中,老年女性患者具有更高的骨骼肌丢失风险[7];邻近70岁的女性(31%)比男性(18.2%)发生肌肉衰减的情况更多[8]。应采用不同标准衡量男女肌肉功能受限的情况[1,9]。
高通量芯片技术一定程度显示与疾病相关的基因变异、表达情况,生物信息学将这些差异表达的基因通过网络分析及其他生物信息分析,如基因本体(gene ontology,GO)分析、通路分析,使蛋白、基因相互作用可视化、系统化,可用于了解疾病发生、发展中复杂的分子机制。
本研究采用公共数据平台的基因芯片数据,通过生物信息学分析,进一步探讨性别与老年肌肉衰减综合征之间的关系。
1 材料与方法
1.1 基因表达谱芯片数据
从美国国立生物技术信息中心(National Center of Biotechnology Information,NCBI)基因表达数据库(Gene Expression Omnibus database,GEO)(https://www.ncbi.nlm.nih.gov/geo/)下载人基因表达芯片数据集GSE8479。基因芯片信息平台为GPL2700,即Sentrix HumanRef-8 Expression BeadChip类型。满足数据集样本共51个,包括25个老年人与26个青年人,骨骼肌组织来源于各自样本股外侧肌,包括老年男性(old male,OM)13例,青年男性(young male,YM)12例,老年女性(old female,OF)12例,青年女性(young female,YF)14例。
1.2 差异基因筛选和样本聚类
将GSE8479芯片数据导入BRB-Array Tools 4.4中进行数据标准化和质量控制。采用中位值将数据标准化。质量控制要求:①截断信号强度大于10,000的值;②基因中位数至少发生1.5倍改变,且不少于20%样本数;③对数化基因表达量变异p<0.01;④数据缺失值≤50%。
基因过滤后,将样本按年龄表型(OM vs.YM、OF vs.YF)筛选差异性表达基因(differentially expressed genes,DEGs)。进行独立样本t检验,DEGs需满足:①p<0.01;②倍数变化>1为上调基因,<1为下调基因。Venny 2.1在线软件分析各组共同的差异基因。根据基因表达情况,判别样本间的距离,采用中心相关和平均距离法进行层级聚类。
1.3 差异基因功能分析和基因集富集分析
采用Cytoscape(http://www.cytoscape.org,Version 3.5.1)插件ClueGo+Cluepedia整合京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路术语,构建DEGs的功能分组通路网络。以KEGG通路术语作为聚类准则,采用双向(富含/缺失)超几何检验,通过Bonferroni校正后,以显著性水平α=0.05查找有影响的基因通路。用Cytoscape插件CytoKegg显示DEGs最显著的通路。
1.4 蛋白质相互作用(protein-protein interaction,PPI)网络构建
将DEGs导入相互作用基因库检索工具(Search Tool for the Retrieval of Interacting Genes,STRING。http://string-db.org)数据库,搜寻已知蛋白质与预测蛋白质之间的相互作用,对蛋白质之间相互作用可能性进行评分,评分>0.4代表两种蛋白间有中度相互作用。进行PPI网络可视化分析。采用Cytoscape进行PPI网络分析;采用其插件MCODE进行子网络聚类分析,得出PPI网络中的相关模块,以MCODE评分>3、节点数>4为标准,得到模块的核心基因。
2 结果
2.1 差异基因筛选和样本聚类
相比YM,OM筛选出DEGs 219个,其中高表达152个,低表达67个;相比YF,OF筛选出DEGs 142个,其中高表达90个,低表达52个;OM和OF共同高表达DEGs 47个,共同低表达21个(图1)。不同性别DEGs层级聚类结果见图1。
2.2 GO分析
OM中,DEGs共涉及9类生物过程,10类分子功能和1类细胞组分。见图2、表1。OF中,DEGs共涉及21类生物过程,5类分子功能,1类细胞组分。见图2、表2(生物过程只列出前10项)。不同性别间,DEGs共涉及1类相同的生物过程,脂肪细胞的分化调节;10类相同的分子功能,蛋白激酶抑制剂活性、蛋白激酶调节剂活性、激酶抑制剂活性和激酶调节剂活性。没有相同的细胞组分。
2.3 KEGG信号通路分析
OM中,DEGs共涉及1条KEGG通路:癌症转录失调。OF中,DEGs共涉及6条KEGG通路:糖酵解/糖异生、FoxO信号通路、胰岛素信号通路、通过细胞色素P450介导的外源化合物代谢、胰高血糖素信号通路、长寿调控通路。见表3。
2.4 PPI网络模块分析
OM网络包括267个节点,1246条边,满足条件的模块4个(图3)。其中评分前三的模块1中心基因为DMD、TPM3、VIM与MYH8,模块2为ZNF415,模块3为CDKN1A(图4)。OF网络包括190个节点,705条边,满足条件的模块7个(图3)。其中评分前四的模块1中心基因为MT1H、MT1X;模块2为DYNLL1、NEDD1,模块3为UCP3,模块4为BGN、LUM(图5)。

图1 各组DEGs数及聚类结果
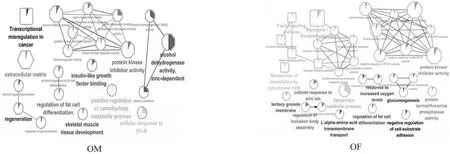
图2 GO及KEGG通路分析结果

图3 DEGs的PPI网络

表1 男性DEGs GO分析结果

图4 男性PPI网络中MCODE评分前三的模块

表2 女性DEGs GO分析结果

图5 女性PPI网络中MCODE评分前四的模块
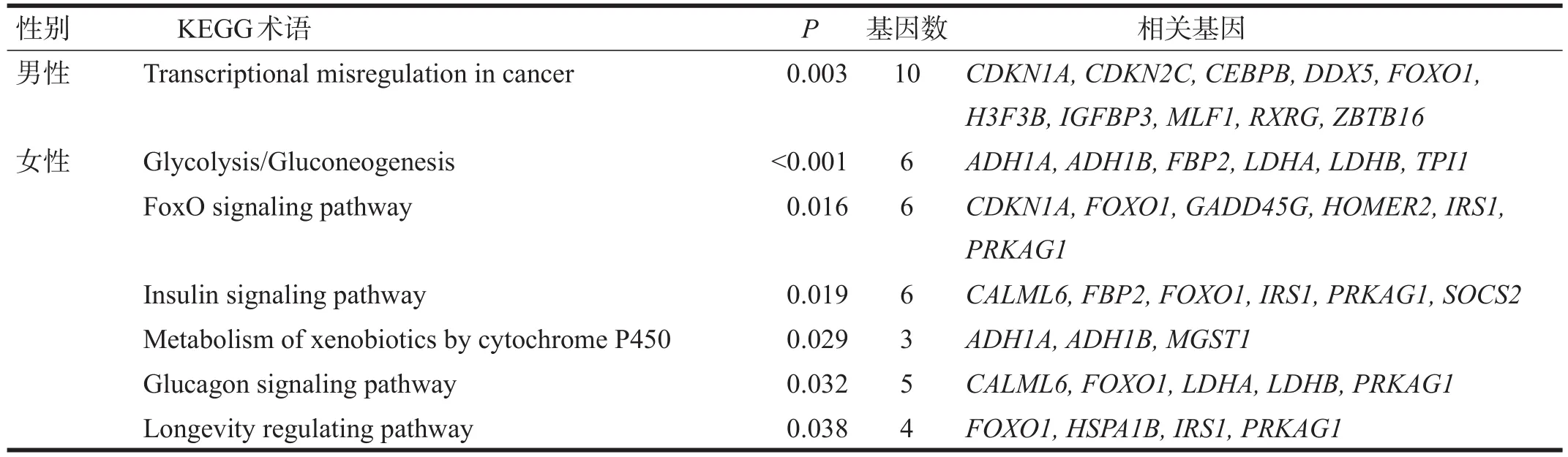
表3 DEGs KEGG通路分析
3 讨论
本研究通过比较老年肌肉衰减综合征患者同正常青年的基因表达,筛选出不同性别的DEGs,发现虽然男性患者DEGs数多于女性,但GO及KEGG通路分析却显示,女性涉及GO类目多于男性。提示性别因素对年龄相关肌肉减少的产生机制存在一定影响:男性进入老年期发生肌肉衰减时,表达变化的基因更为集中;而女性涉及的生物机制可能更复杂。
骨骼肌作为人体最大的外周胰岛素靶器官[10],约消耗80%血糖,并参与一些化合物的代谢;随着衰老出现的骨骼肌衰减或萎缩可导致IR[11-12]。本研究显示,男女老人部分DEGs集中于脂肪细胞分化的生物过程中,并全部高表达。研究表明,肌萎缩[13]与IR[10]可单独或共同造成脂肪异位沉积,而成肌细胞在一定条件下还可向脂肪细胞分化[14],提示肥胖与骨骼肌衰减有密切联系。
DEGs富集模块还有蛋白激酶抑制剂、调节剂活性,并同样呈高表达。蛋白激酶参与多种细胞功能,可促进细胞增殖及对胰岛素应答,并抑制细胞凋亡;抑制剂高表达会抑制蛋白激酶,进而影响正常细胞生长。如在衰老的肌肉组织中,细胞周期素依赖的蛋白激酶抑制剂1A(cyclin-dependent kinase inhibitor 1A,CDKN1A)活性增强,使静止期肌卫星细胞G0或G2期阻滞,导致静止期细胞向活化细胞转换,再生及自我更新能力下降[15]。针对蛋白激酶抑制剂表达和功能的调节或许是治疗年龄相关肌肉丢失的潜在靶点[16]。
PPI网络和模块分析可判断一个蛋白在该网络中的地位[17]。本研究显示,老年男性抗肌萎缩蛋白(dystrophin,DMD)、波形蛋白(vimentin,VIM)、原肌球蛋白α-3链(tropomyosin α-3,TPM3)是重要的节点。DMD基因是人体最大的基因之一。动物研究发现[18],老年大鼠肌肉量丢失前期,该基因表达就已经上调,而编码的蛋白表达量下调。本研究结果与之一致。TPM3是一种结合于肌动蛋白的蛋白质,VIM则是一种中间纤维蛋白,通常被认为是肌细胞再生指标[19],两者均参与细胞骨架的构成[20]。值得注意的是,VIM的高表达还与前列腺癌有密切联系[21-22],可能预示男性肌肉衰减综合征与前列腺癌发病之间有某种联系。
在老年女性中,金属硫蛋白(metallothionein,MT)、动力蛋白轻链(dynein light chain,DYNLL)是重要的节点。MT是一种富含半胱氨酸的低分子量蛋白质,具有能与金属离子结合的位点,参与调节机体内金属离子代谢,尤其是锌稳态的过程[23],对抗氧化应激反应。MT1H是其4种亚型的一种,可作为诊断与治疗女性乳腺癌转移的新靶点[24]。提示女性肌肉衰减综合征与乳腺癌有关。DYNLL1属动力蛋白家族,负责转运蛋白[25];它在肌肉细胞池的富集可影响肌肉细胞长度及细胞核位置[26],而其变异与神经系统疾病如脊髓侧索硬化症[27]、阿尔茨海默病[28]等有密切联系。有研究显示[29],动力蛋白与促凋亡的caspase-3在衰老小鼠神经肌肉接头处高表达,凋亡因子向神经元胞体逆行转运。提示DYNLL1可能是神经退行性病变与老年骨骼肌衰减症共同靶点之一。
综上所述,本研究通过生物信息学方法,挖掘与老年肌肉衰减综合征发生相关的DEGs,并探讨其性别差异,结果提示,老年肌肉衰减综合征的发病机制可能存在性别差异,一些特定生物学过程可能与老年肌肉衰减综合征相关;某些基因,如VIM、MT1H,可能预示着肌肉衰减综合征还伴随发生其他不良事件。
由于高通量芯片技术结果较多,统计方法存在局限,本研究结果需要进一步实验支持。
[1]Chen LK,Liu LK,Woo J,et al.Sarcopenia in Asia:consensus report of the Asian Working Group for Sarcopenia[J].J Am Med DirAssoc,2014,15(2):95-101.
[2]Cruz-Jentoft AJ,Baeyens JP,Bauer JM,et al.Sarcopenia:European consensus on definition and diagnosis:Report of the European Working Group on Sarcopenia in Older People[J].Age Ageing,2010,39(4):412-423.
[3]Liu LK,Lee WJ,Liu CL,et al.Age-related skeletal muscle mass loss and physical performance in Taiwan:implications to diagnostic strategy of sarcopenia in Asia[J].Geriatr Gerontol Int,2013,13(4):964-971.
[4]Chen LK,Lee WJ,Peng LN,et al.Recent advances in sarcopenia research in Asia:2016 Update from the Asian Working Group for Sarcopenia[J].J Am Med Dir Assoc,2016,17(8):e761-e767.
[5]Fielding RA,Vellas B,Evans WJ,et al.Sarcopenia:an undiagnosed condition in older adults.Current consensus definition:prevalence,etiology,and consequences.International Working Group on Sarcopenia[J].J Am Med Dir Assoc,2011,12(4):249-256.
[6]Pedersen BK,Febbraio MA.Muscles,exercise and obesity:skeletal muscle as a secretory organ[J].Nat Rev Endocrinol,2012,8(8):457-465.
[7]Park SW,Goodpaster BH,Lee JS,et al.Excessive loss of skele-tal muscle mass in older adults with type 2 diabetes[J].Diabetes Care,2009,32(11):1993-1997.
[8]Kirchengast S,Huber J.Gender and age differences in lean soft tissue mass and sarcopenia among healthy elderly[J].AnthropolAnz,2009,67(2):139-151.
[9]Kim JH,Choi SH,Lim S,et al.Sarcopenia and obesity:gender-different relationship with functional limitation in older persons[J].J Korean Med Sci,2013,28(7):1041-1047.
[10]Kitessa SM,Abeywardena MY.Lipid-induced insulin resistance in skeletal muscle:the chase for the culprit goes from total intramuscular fat to lipid intermediates,and finally to species of lipid intermediates[J].Nutrients,2016,8(8):E466.
[11]Roubenoff R.Sarcopenic obesity:the confluence of two epidemics[J].Obes Res,2004,12(6):887-888.
[12]Lee SW,Youm Y,Lee WJ,et al.Appendicular skeletal muscle mass and insulin resistance in an elderly Korean population:the Korean social life,health and aging project-health examination cohort[J].Diabetes Metab J,2015,39(1):37-45.
[13]Wiggs MP.Can endurance exercise preconditioning prevention disuse muscle atrophy?[J].Front Physiol,2015,6(63):1-15.
[14]Zhang F,Deng B,Wen J,et al.PPAR gamma and MyoD are differentially regulated by myostatin in adipose-derived stem cells and muscle satellite cells[J].Biochem Biophys Res Commun,2015,458(2):375-380.
[15]Sousa-Victor P,Gutarra S,Garcia-Prat L,et al.Geriatric muscle stem cells switch reversible quiescence into senescence[J].Nature,2014,506(7488):316-321.
[16]Mohan A,Asakura A.CDK inhibitors for muscle stem cell differentiation and self-renewal[J].J Phys Fit Sports Med,2017,6(2):65-74.
[17]Callaway DS,Newman ME,Strogatz SH,et al.Network robustness and fragility:percolation on random graphs[J].Phys Rev Lett,2000,85(25):5468-5471.
[18]Hughes DC,Marcotte GR,Marshall AG,et al.Age-related differences in dystrophin:impact on force transfer proteins,membrane integrity,and neuromuscular junction stability[J].J GerontolABiol Sci Med Sci,2017,72(5):640-648.
[19]Vater R,Cullen MJ,Harris JB.The expression of vimentin in satellite cells of regenerating skeletal muscle in vivo[J].Histochem J,1994,26(12):916-928.
[20]Holland A,Henry M,Meleady P,et al.Comparative label-free mass spectrometric analysis of mildly versus severely affected mdx mouse skeletal muscles identifies annexin,lamin,and vimentin as universal dystrophic markers[J].Molecules,2015,20(6):11317-11344.
[21]魏军成.波形蛋白对前列腺癌细胞侵袭与转移的影响[J].癌症,2008,27(1):30-34.
[22]Vyas AR,Singh SV.Functional relevance of D,L-sulforaphane-mediated induction of vimentin and plasminogen activator inhibitor-1 in human prostate cancer cells[J].Eur J Nutr,2014,53(3):843-852.
[23]Dong G,Chen H,Qi M,et al.Balance between metallothionein and metal response element binding transcription factor 1 is mediated by zinc ions(review)[J].Mol Med Rep,2015,11(3):1582-1586.
[24]Tai SK,Tan OJ,Chow VT,et al.Differential expression of metallothionein 1 and 2 isoforms in breast cancer lines with different invasive potential:identification of a novel nonsilent metallothionein-1H mutant variant[J].Am J Pathol,2003,163(5):2009-2019.
[25]Kreko-Pierce T,Eaton BA.The Drosophila LC8 homolog cut up specifies the axonal transport of proteasomes[J].J Cell Sci,2017,130(19):3388-3398.
[26]Folker ES,Schulman VK,Baylies MK.Muscle length and myonuclear position are independently regulated by distinct dynein pathways[J].Development,2012,139(20):3827-3837.
[27]Xie Y,Zhou B,Lin MY,et al.Endolysosomal deficits augment mitochondria pathology in spinal motor neurons of asymptomatic fALS mice[J].Neuron,2015,87(2):355-370.
[28]Kimura N,Samura E,Suzuki K,et al.Dynein dysfunction reproduces age-dependent retromer deficiency:concomitant disruption of retrograde trafficking is required for alteration in beta-amyloid precursor protein metabolism[J].Am J Pathol,2016,186(7):1952-1966.
[29]Garcia ML,Fernandez A,Solas MT.Mitochondria,motor neurons and aging[J].J Neurol Sci,2013,330(1-2):18-26.
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
基层中医药(2022年4期)2022-07-22
体育科技文献通报(2022年3期)2022-05-23
医学综述(2021年16期)2021-12-01
中医眼耳鼻喉杂志(2021年2期)2021-07-21
科学(2020年2期)2020-08-24
儿童故事画报(2020年12期)2020-06-23
三农资讯半月报(2020年3期)2020-03-09
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
运动(2018年14期)2018-07-16
