
破壁预处理对小球藻藻渣产沼气性能的影响
2018-03-30 06:52周银存赵新仕毕生雷胡守珍张乃群
中国沼气 2018年1期
周银存, 赵新仕, 杨 迪, 毕生雷, 鲁 龙, 胡守珍,, 张乃群
(1.南阳师范学院 生命科学与技术学院, 河南 南阳 473061; 2.河南天冠企业集团有限公司 车用生物燃料技术国家重点实验室, 河南 南阳 473000)
小球藻是一种普生性单细胞绿藻[1],适应性比较强、广泛分布于淡水、海水等各种环境,甚至冰层、火山口等极端环境也有分布。小球藻因其生长迅速、生长周期短、占地面积小、可以使用工业化设备进行规模化培养等特点而备受关注[2],小球藻用途广泛,富含的藻多糖、藻蛋白、天然色素、小球藻生长因子等高附加值产物被使用在食品、美容等领域,高油脂含量也决定了其是制备生物柴油的优质原料[3]。
虽然小球藻可利用价值高,但小球藻产品多为胞内产物。小球藻细胞壁相对较厚且细胞组成架构复杂[4],只有细胞破碎才能获得细胞内部的高附加值产物。不同的细胞破碎工艺对小球藻造成的破坏程度也不同,直接影响到目的产物的提取效率和产量。同时,不同的小球藻细胞破碎工艺会出现环保、能耗等不同的问题,因此需要根据小球藻产品来选择合适的细胞破碎工艺。
小球藻细胞进行高附加值产物提取后的残渣仍然含有丰富的糖类、蛋白类等营养物质,直接舍弃不仅污染环境还会造成不必要的浪费,而沼气发酵能够有效降解其它发酵过程不能利用的富余营养物质。近年来,各类大中型沼气工程的建设,实现了厨余垃圾、秸秆等废弃物的降解,不仅改善了生态环境,还生产了大量沼气这一清洁能源为民所用,是进行生态环境重建的重要举措[5]。通过沼气发酵进一步利用小球藻的废液和藻渣,已经成为科研人员探索的一种新思路。本文通过对异养小球藻藻渣产沼气工艺进行研究,对比探讨不同的小球藻破壁方式对藻渣沼气生产的分解阶段和产沼气能力的影响,从而获得藻渣产沼气的最佳预处理工艺参数。
1 材料与方法
1.1 试验材料
异养小球藻:异养培养的原始小球藻(Chlorellaprotothecoides),由河南天冠集团车用生物燃料国家重点实验室提供,细胞内油脂含量高达50%以上;
异养小球藻培养液:取自车用生物燃料国家重点实验室,原始小球藻质量体积浓度为120 g·L-1;
异养小球藻采用各种提取方法提取后的藻液:取自车用生物燃料国家重点实验室,存于展示柜中备用;
接种用污泥:取自天冠集团技术中心,TS为1%,在水浴锅中50 ℃活化1 d备用;
其它试验材料包括玻璃器皿、试剂等均外购。
1.2 试验设备
厌氧发酵系统:自制,主要由恒温水浴锅、发酵罐、集气瓶和集水瓶组成。发酵罐、集气瓶、集水瓶均为l L广口瓶,采用胶塞密封,发酵罐置于恒温水浴锅中,发酵罐口设料液取样口及导气管,导气管连接集气瓶和集水瓶,具体见图1。
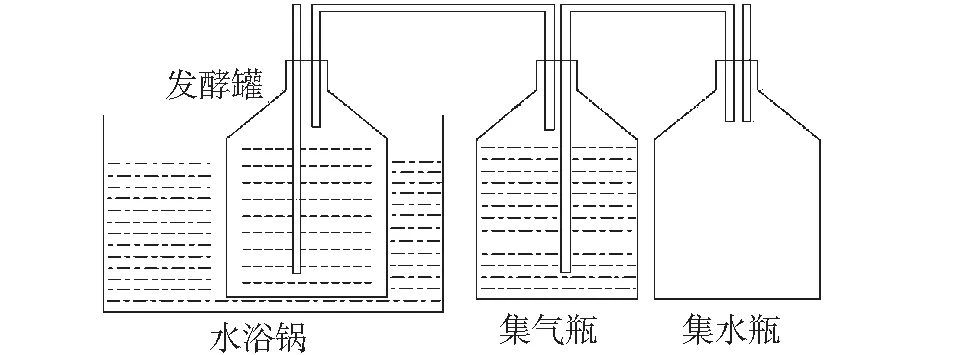
图1 沼气发酵装置示意图
沼气发酵和预处理工艺中所用设备均为外购,主要有上海精密DK-600型恒温水浴锅、上海皓庄LNB2.5-10F型马弗炉、梅特勒FE20-FiveEasy Plus型pH计,上海博珍202-2型电热恒温干燥箱,深圳昌鸿QCOD-3E型COD测定仪,深圳昌鸿NH-6N型氨氮测定仪,长沙湘仪L400型离心机,瑞士步琦R-300型旋转蒸发仪,安捷伦2100型液相色谱仪,武汉四方光电科技Gasboard-3200P 沼气分析仪等。
1.3 试验方法
1.3.1 原料制备方法
培养液藻渣[6]:取异养小球藻培养液藻渣,藻渣油脂含量51.3%。
球磨破碎提油后残渣[7]:异养小球藻培养液使用球磨法进行细胞破碎、提油后余藻渣备用,藻渣油脂含量7.9%。
酶解提油后残渣[8]:异养小球藻培养液使用酶解法进行细胞破碎、提油后余藻渣备用,藻渣油脂含量25.8%。
酸热法破碎提油后残渣[9]:异养小球藻培养液使用酸热法进行细胞破碎、提油后余藻渣备用,藻渣油脂含量0.1%。
酯化提柴油后残渣[10]:异养小球藻培养液原位转酯化法制备生物柴油后余藻渣备用,藻渣油脂含量0.1%。
1.3.2 原料处理方法
异养小球藻培养液及采用各种提取方法提油后的藻液,使用离心机离心制成藻渣后,取10 g置于烘箱中测100 ℃干重;取藻渣置于广口瓶中,加入接种用污泥,配成干重为10%的沼气发酵液。
1.3.3 沼气发酵方法
取1000 mL沼气发酵液置于发酵罐中,沼气发酵温度为50 ℃,每天摇动发酵罐,每周取样送检并补充集气瓶中水,使用排水法计算排气量[11],其中一个发酵系统连续两周停止产气后,整个试验停止。
1.3.4 检测方法
排气量:直接用量筒量取集水瓶中水的体积;pH值:直接使用pH计检测;挥发性脂肪酸:使用液相色谱仪检测乙酸、丙酸等成分含量,然后加总;碳源:使用液相色谱仪检测葡萄糖、木糖等成份含量,然后加总;氨氮:使用氨氮检测仪检测;COD:使用COD检测仪检测;甲烷含量:使用Gasboard-3200P 沼气分析仪检测。
1.3.5 计算公式
计算公式如下:
干重(g·L-1)=[藻渣烘干后重量(g)/藻渣体积(mL)]×100
2 试验结果
2.1 不同原料发酵体系pH值变化情况
使用不同预处理工艺原料各发酵体系的pH值变化情况如图2。
从图2可以看到,随着发酵的进行,5个发酵体系中,使用球磨法、酯化法、酶解液和培养液为原料的4个发酵体系的pH值快速下降,在达到稳定状态后又小幅回升,而使用酸热法原料的发酵体系则一直处于下降趋势中。
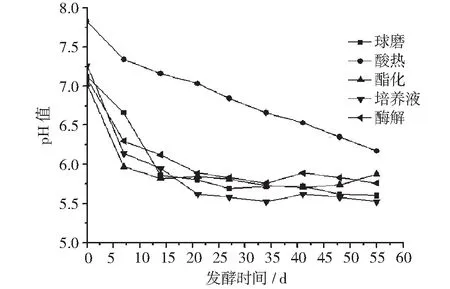
图2 pH值变化图
2.2 不同原料发酵体系变化情况
在使用不同预处理工艺原料的发酵体系中,氮的平衡是非常重要的因素[12],不同预处理工艺产生的残渣由于细胞破碎程度不同,可利用程度也差异较大。小球藻残渣的初始溶解性有机氮的含量及在发酵过程中的变化趋势详见图3。
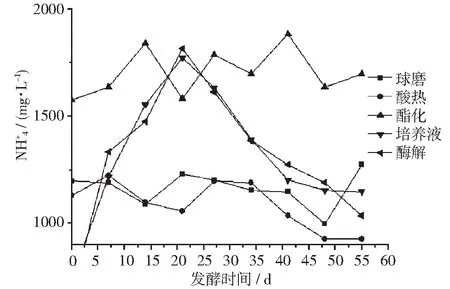
图 变化图
从图3可以看出,酯化制柴油后的藻渣作为原料时,其氨氮一直处于较高的水平,培养液、酶解原料最初氨氮为0,然后快速上升,20 d后又迅速下降,球磨、酸热原料氨氮一直处于较低的水平。
2.3 不同原料发酵体系COD变化情况
COD即化学需氧量[13],是沼气发酵中的重要参数,使用不同预处理工艺原料的发酵体系中,其COD变化情况如图4。
从图4可以看出,COD的整体变化趋势是先降低,再升高,然后再降低。其中,球磨法原料所在的发酵体系COD一直处于较高的水平,且在发酵的前48 h远远高于其它原料所在发酵体系。而其它原料所在的发酵体系COD一直处于较低的水平,在300000 mg·L-1以下波动。发酵结束后5种原料的发酵体系COD均高于150000 mg·L-1。

图4 COD变化图
2.4 不同原料发酵体系总碳含量变化情况
不同预处理工艺原料的发酵体系中的总碳含量的变化情况详见图5。

图5 总碳变化图
从图5可以看出,酸热法原料所在的发酵体系中总碳含量一直处于较高的水平,而培养液、酶解法原料所在的发酵体系中总碳含量先快速上升,然后又快速下降,酯化法、球磨法原料所在的发酵体系中总碳含量一直处于较低的水平。
2.5 不同原料发酵体系VFA含量变化情况
使用不同预处理工艺原料的发酵体系中,其可挥发性脂肪酸含量的变化情况详见图6。
从图6可以看出,球磨法、酸热法、酯化法原料所在的发酵体系中VFA一直处于较低的水平,且比较平衡,而培养液、酶解液所获得原料所在的发酵体系中VFA先快速升高,又下降,但整个发酵过程中培养液、酶解法所获得原料的VFA含量一直高于其它原料所在的发酵体系。
2.6 不同原料发酵体系累积产气量变化情况
沼气是沼气发酵的最终目的,产气量的高低反映了沼气发酵体系的生产能力,以及原料的利用能力。采用不同预处理工艺原料的发酵体系,其产气量的变化情况详见图7。
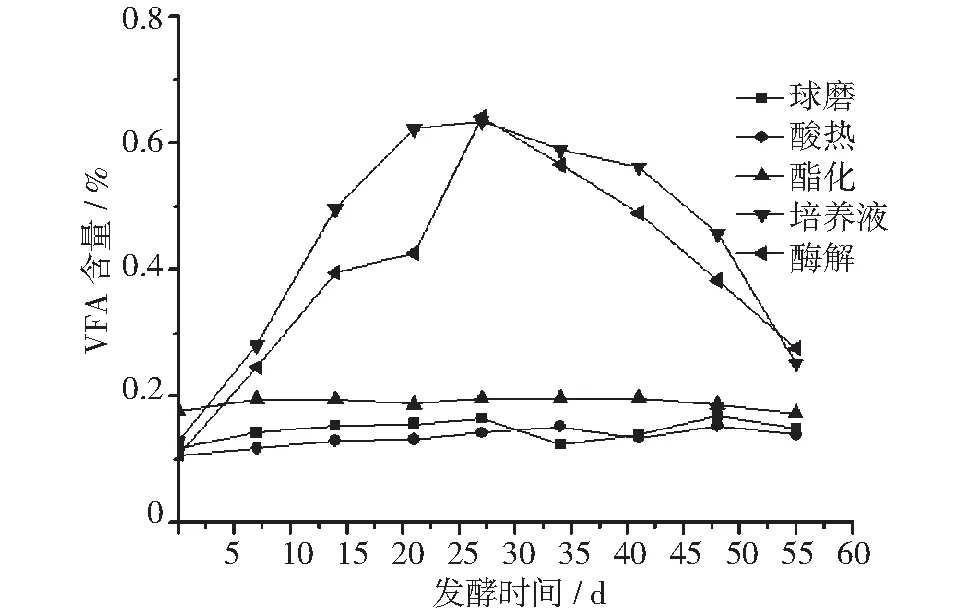
图6 VFA变化图
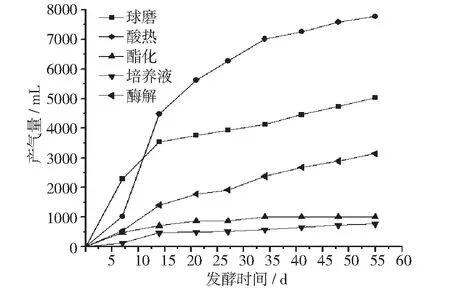
图7 累积产气量变化图
从图7可以看出,酸热法原料所在的发酵体系产气量最高,接近8000 mL,然后依次是球磨法、酶解法、酯化法、培养液。培养液、酶解法原料所在的发酵体系产气量最低,不足1000 mL。球磨法、酯化法原料所在的发酵体系产气量处于中间水平。
2.7 不同预处理工艺的发酵体系所产沼气中 CH4含量的变化情况
试验通过 Gasboard-3200P 沼气分析仪对甲烷含量进行测定[14],精确度为0.1%。试验分别在发酵的第14,27,41 天测CH4含量,具体数据如表1所示。总体来看,各原料所产沼气中的CH4含量均随着发酵时间的延长而升高,说明发酵原料逐步分解,步入了甲烷化阶段,此阶段的甲烷菌活性较强。酸热法所产的沼气CH4含量是最高的,第41天高达70.10%,平均甲烷含量为57.20%。
3 分析与讨论
3.1 不同预处理工艺对pH值的影响
结构复杂的大分子有机质无法直接被沼气细菌利用,需要经过水解作用分解成小分子有机质,然后进一步经过酸化作用分解成沼气细菌能够利用的短链有机酸。

表1 不同预处理工艺的发酵体系所产沼气中 CH4含量的比较 (%)
但水解过程、酸化过程和产沼气过程需要均衡进行。如果水解过程和酸化过程超过了产沼气过程的反应速度,就会导致产酸过量,从而引起pH值降低,沼气菌就需要更多能量从细胞体内向体外提供质子以维持其细胞质的生长环境[15]。因此有必要了解pH值的变化规律,以提高沼气发酵效率。
从图2的pH值变化中可以看到,随着发酵的进行,使用球磨法提油、酯化法制柴油、酶解液提油获得的3种原料和培养液原料4个发酵体系的pH值首先快速下降,在达到稳定状态后又小幅回升。而使用酸热法提油原料的发酵体系则一直处于稳步下降趋势中。
由于预处理工艺对原料结构造成不同的影响,球磨法、酯化法、酶解法3种原料的细胞受到破坏程度有限,与培养液原料差别不大。在发酵初期,受到破坏并成为细胞碎片的部分能够被水解菌和产酸菌利用,从而使水解菌和产酸菌活跃,加速了水解、酸化进程,反映在pH值上就是pH值快速下降,而在水解过程和酸化过程将能够快速利用的有机质利用完成后,藻细胞需要慢慢被分解,从而维持pH值平衡状态。随后短链有机酸被逐渐消耗,pH值缓慢上升。而对于酸热法获得的原料,由于细胞破坏完全,原料基本都是细胞碎片[9],因此整个沼气发酵过程的水解和产酸都比较旺盛,从而使pH值一直处于稳步的下降状态。
3.2 不同预处理工艺对的影响
对于固体有机物,沼气发酵体系首先要将其分解为能溶于水、能够被酸化细菌利用的小分子有机质,因此,溶解性有机氮的变化趋势能够反映沼气发酵体系对底物的利用情况[11]。

3.3 不同预处理工艺对COD的影响
COD是指处理样品时,所消耗的强氧化剂的数量,代表了样品中能够被氧化的有机物质数量。通常情况下,小分子有机物能够被完全氧化,大块的复杂有机物由于难以被强氧化剂完全破坏只是被部分氧化。能够被氧化的有机物通常能够被沼气发酵细菌利用,因此COD可以反映反应体系中底物浓度的高低。
从图4的COD变化中可以看出,COD的整体变化趋势是先降低后升高,然后再降低,说明随着底物被分解、酸化、产甲烷这一过程,底物浓度降低,从而引起COD有所降低。而水解细菌的活动又不断将不能利用的底物进一步分解成小分子有机物,从而导致COD又有所升高。但分解、酸化、产甲烷始终是一个动态的平衡的体系,因此COD的变化趋势虽然波动较大,但一直处于一个平衡的区间。
在5种原料中,球磨提油后原料所在的发酵体系COD一直处于较高的水平,说明球磨后形成的细胞碎片能够不断被分解。而培养液、酶解液原料所在的发酵体系COD一直处于较低的水平,说明小球藻细胞难以被分解。酸热法、酯化法获得的两种原料,由于细胞在强硫酸作用下已经被破坏,细胞结构中的淀粉、蛋白质被强酸在高温下碳化无法再通过沼气发酵水解出更多营养物质,所以其所在发酵体系的COD也一直处于较低的水平。
发酵结束后,五种原料的发酵体系COD均高于150000 mg·L-1,这可能是未被利用的细胞以及在提油时残留的有机溶剂,说明发酵体系中留存的细胞碎片以及其它有机溶剂虽然影响了COD,但不能被沼气细菌利用。
3.4 不同预处理工艺对总碳含量的影响
复杂的大块的有机物难以被沼气细菌直接利用,在分解过程中被水解菌群分解为能够被酸化细菌利用的糖类、醇类,总碳含量反映了糖类和醇类的总和,其含量高低能够说明分解过程进展情况。
从图5的总碳含量变化中可以看出,酸热法获得原料所在的发酵体系中总碳含量一直处于较高的水平,说明酸热法对小球藻细胞破坏比较彻底,能够被水解细菌彻底利用。
培养液、酶解法获得原料所在的发酵体系中总碳含量先快速上升,然后又快速下降,说明培养液、酶解法所获得原料不能被水解细菌快速分解,其利用有一定过程。在此过程中酸化细菌不能获得足够的营养物质而活性受限,随着分解过程的进行,酸化细菌能够获得的营养物质越来越多,活性增强,逐步超过水解细菌分解能力,导致总碳含量快速下降。
酯化法、球磨法获得原料所在的发酵体系中总碳含量一直处于较低的水平,说明水解细菌、酸化细菌在发酵过程中形成了动态平衡,发酵体系比较稳定,但这两种原料通过分解获得的营养物质偏少。
3.5 不同预处理工艺对VFA含量的影响
酸化细菌利用分解过程中产生的小分子有机物,从而产生了乙酸、丙酸、丁酸等可挥发性脂肪酸,这些短链脂肪酸能够被甲烷细菌直接利用从而产生沼气。VFA含量可以代表乙酸、丙酸等可挥发性脂肪酸含量的多少,也能够说明酸化过程进展情况[17]。
从图6的VFA变化中可以看出,球磨法、酸热法、酯化法所获得原料所在的发酵体系中VFA一直处于较低的水平,且比较平衡,说明酸化过程、产甲烷过程形成了动态平衡,沼气发酵比较稳定。
相对于其它原料反应体系,培养液、酶解液所获得原料所在的发酵体系中VFA先快速升高,又下降,结合图5,可以说明这两种原料的细胞壁受到破坏程度低、不能快速被分解细菌直接利用,因此酸化细菌前期受到抑制,产酸较少。但随着分解过程的进行,仍有部分大分子有机物被分解成能够被酸化细菌利用的营养物质,从而导致酸化细菌活性增强,产酸快速增加,引起VFA含量的提高。只是分解过程较为缓慢,不能满足快速增加的酸化细菌营养需求,导致产酸减少,VFA含量降低。
3.6 不同预处理工艺对产气量的影响
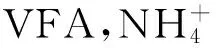
从图7的产气量变化中可以看出,酸热法获得原料所在的发酵体系产气量最高,接近8000 mL,然后依次是球磨法、酶解法、酯化法、培养液。
4 小结
[1] 鲁 龙,毕生雷,金洪波,等.强酸破碎小球藻细胞壁工艺条件的优化[J].食品与发酵科技,2016, 52 (2): 39-42.
[2] Jin-Young An, Sang-Jun Sim, Jin Suk Lee. et al. Hydrocarbon production from secondarily treated piggery wastewater by the green alga Botryococcus braunii[J].Journal of Applied Phycology,2003, 15 (2): 185-191.
[3] 毕生雷,张成明,郑世文,等.异养小球藻原料直接制备生物柴油的研究[J].中国油脂,2016, 41 (10): 48-52.
[4] 钟韵山,徐仰仓,荆柏林,等.小球藻破壁技术研究进展[J].食品研究与开发,2014,35 (14): 120-124.
[5] 乔 慧,陈 灏,赵玉柱. 沼液处理液养殖普通小球藻的试验研究[J].中国沼气,2016, 34(6):115-120.
[6] 毕生雷,张成明,李十中,等.异养小球藻半连续发酵生产油脂工艺探讨[J].食品与发酵科技,2014(5):36-40,48.
[7] 鲁龙,毕生雷,金洪波,吴娟,张鹏飞. 研磨法破碎小球藻细胞工艺优化[J].中国酿造,2017(1):116-119.
[8] 邢欢,许文宗,张志荣,钟韵山,李婕,徐仰仓. 绿色木霉对小球藻细胞壁的酶解作用[J].微生物学通报,2015(6):975-980.
[9] 鲁龙,毕生雷,金洪波,吴娟,张鹏飞. 强酸破碎小球藻细胞壁工艺条件的优化[J].食品与发酵科技,2016(2):39-42,47.
[10] 毕生雷,张成明,郑世文,金洪波,吴娟,鲁龙,张鹏飞. 异养小球藻原料直接制备生物柴油的研究[J].中国油脂,2016(10):48-52.
[11] 高 鑫. 秸秆类原料厌氧发酵产沼气试验研究[D].上海:华中农业大学,2013.
[12] 张 波,徐剑波,蔡伟民. 有机废物厌氧消化过程中氨氮的抑制性影响[J].中国沼气,2003,21(3):26-31.
[13] 叶小梅,常志州,钱玉婷,等.江苏省大中型沼气工程调查及沼液生物学特性研究[J].农业工程学报,2012,28(6):222-227.
[14] 王晓曼. 早熟禾厌氧发酵特性和产气潜力的研究[D].西北农林科技大学,2010.
[15] 郭萃萍.稻秆干发酵制沼气工艺参数的研究[D].石家庄:河南农业大学,2012.
[16] 许之扬. 餐厨垃圾固态厌氧消化过程内源性抑制效应研究[D].无锡:江南大学,2014.
[17] 毕少杰,孙 宇,孙志远,等.酸化处理对牛粪厌氧发酵有机酸和细菌多样性的影响[J].中国沼气,2015(4):18-25.
猜你喜欢
南方农业(2022年12期)2022-11-23
西南农业学报(2022年8期)2022-10-28
科学技术创新(2022年28期)2022-10-21
中学生物学(2021年8期)2021-11-02
考试与评价·高二版(2021年2期)2021-09-10
海洋通报(2020年3期)2020-10-20
可再生能源(2020年8期)2020-08-17
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
陶瓷学报(2019年5期)2019-01-12
中国果业信息(2019年1期)2019-01-05
