miR-17调控自噬保护高糖环境下内皮细胞*
2018-03-15 03:02李罗成王志维吴智勇胡知朋阮永乐李博文
微循环学杂志 2018年1期
李罗成 王志维 吴智勇 胡知朋 阮永乐 李博文
血管内皮功能失调和损伤在糖尿病血管病变发生发展中起着关键作用,也是糖尿病引起靶器官损害的始动环节。长期高血糖诱发的氧化应激反应是糖尿病血管内皮损伤的重要因素[1]。减轻甚或阻断糖尿病引起的血管内皮损伤有利于防治糖尿病并发症,但在这一临床难题上尚缺少确实有效的手段。
微小RNA(microRNA,miRNA)是一种非编码RNA,作为一种基因表达的转录后调节因子,miRNA可通过多种信号途径调控靶细胞mRNA的稳定性、翻译效率和生物学功能,包括细胞对氧化还原过程的反应[2]。机体细胞处于氧化应激状态时,会激活自噬清除细胞内错误折叠的蛋白、损伤的DNA和线粒体等,以利于细胞存活。研究表明,与正常人相比,糖尿病患者血清miR-17表达显著减少[3],而miR-17在调控细胞自噬中起着重要作用[4]。miR-17是否参与内皮细胞自噬的调控并影响内皮细胞的氧化应激反应,值得深入探讨。本研究拟通过体外培养人脐静脉内皮细胞(Human Umbilical Vein Endothelial Cells, HUVECs)并加入不同浓度葡萄糖干预,观察miR-17调控的自噬在抗内皮细胞氧
化应激和凋亡中的作用。
1 材料与方法
1.1 主要材料和试剂
HUVECs购于Lonza公司(瑞士),兔抗LC3B(批号:ab48394)、Beclin-1(批号:ab55878)、β-actin(批号:ab8226)多克隆抗体和辣根过氧化物酶(HRP)标记羊抗兔IgG(批号:ab6721)购自英国Abcam公司,兔抗sequestosome 1(SQSTM1,批号:sc-25575)购自美国Santa Cruz生物技术公司,3-甲基腺嘌呤(3-Methyladenine, 3-MA, 批号:HY-19312)购自美国MedChemExpress公司, 超氧化物歧化酶(Superoxide Dismutase, SOD)活性检测试剂盒(批号:S0101)、脂质氧化丙二醛(Malonaldehyde , MDA)检测试剂盒(批号:S0131)、RIPA 裂解液(批号:P0013C)、BCA蛋白检测试剂盒(批号:P0010S)购自碧云天生物技术有限公司,miRNA提取试剂盒(批号:SNC50)购自美国Sigma-Aldrich公司,miR-17 mimics 、miR-Control mimics和PCR引物由广州市锐博生物科技有限公司(ribobio)合成,见表1。

表1 miR-17 mimics、miR-control mimics及引物序列
1.2 方法
1.2.1HUVECs培养及传代:HUVECs复苏后,采用含10%胎牛血清和1%双抗(青链霉素混合液)的RPMI 1640 培养液,置于37℃、5% CO2的培养箱中培养,2-3天更换培养液。通过倒置相差显微镜观察细胞,当细胞生长面积达到培养板底部的80%或出现接触抑制时,进行传代。选择传代次数为3-7次的HUVECs用于以下实验。
1.2.2HUVECs转染miR-17 mimics:转染前一天将HUVECs按50%的融合度接种到24孔板,并于每个孔内加入500μl含10% FBS的无抗生素 RPMI 1640培养基。将2μl Lipofectamine®2000用无血清培养基opti-MEM稀释至50μl,混合均匀后在室温下孵育5min。再分别于50μl opti-MEM中加入miR-17 mimics (工作浓度50nmol/L)、miR-control mimics(工作浓度50nmol/L),将两种液体分别与稀释的Lipofectamine®2000混匀,在室温下孵育20min,然后加入到24孔板的每个孔中,用opti-MEM I Reduced Serum Medium将总体积补足至500μl并混匀。干预实验设非转染组(未转染miR-17 mimics)、miR-17 mimics组(转染miR-17 mimics)和miR-17 mimics+3-MA组(转染miR-17 mimics并加入自噬抑制剂3-MA,3-MA工作浓度2mmol/L,转染24h后加入),于37℃培养箱内培养、转染24h,更换培养基后每组分别加入不同浓度葡萄糖(对照组:5.5mmol/L;高糖组:16.7mmol/L),干预48h后进行细胞收集和检测分析,每个样品设置6个复孔。
1.2.3qRT-PCR检测miR-17 mimics转染效率:收集转染24 h后的HUVECs,提取miRNA进行反转录,miRNA的提取根据miRNA提取试剂盒(mirPremierRMiRNA Isolation Kit)的说明书操作方法进行。反转录体系如下:RNA引物组合12μl,5×RT 反应缓冲液5μl,25mmol/L dNTPs 1μl,25U/μl RNA酶抑制剂1μl,200U/μl M-MLV Rtase 1μl,Oligo(dt)18 μl,无DNA酶双蒸水4μl,总体积25μl。合成cDNA并以其为模板扩增,扩增体系如下:SYBR Green Mix 12.5μl,F-引物 0.5μl,R-引物0.5μl,双蒸水9.5μl,cDNA模板2μl,总体积25μl。qRT-PCR反应程序:预变性95℃ 20s,变性95℃ 15s;退火60℃ 20s,共40个循环。内参为U6,PCR反应在96板中完成,每个样本设3个复孔。通过PCR仪的自带软件(ABI Prism 7300 SDS Software)进行数据分析。根据公式用CT值计算基因的起始模板浓度(ITC),ITC=2-△△CT。△△CT的计算方法为△CT(n)=CT目的基因(n)-CT内参基因(n),△△CT(n)=△CT(n)-△CT(1)。
1.2.4Western Blotting检测LC3II/LC3I、SQSTM1、Beclin-1表达:收集各干预条件下培养48h的HUVECs,加入适量RIPA细胞裂解液,提取总蛋白,按BCA蛋白检测试剂盒说明书进行操作,测定蛋白浓度、绘制标准曲线并计算样品蛋白浓度值。根据蛋白定量结果,取50μg蛋白质经15%聚丙烯酰胺凝胶电泳,半干法转移到PVDF膜。转膜后用含5%脱脂奶粉的TBST进行封闭,再加入一抗(稀释比例LC3B, 1∶1 000; Beclin1, 1∶1 000; SQSTM 1, 1∶1 000; β-actin, 1∶1 500)室温孵育2h,TBST洗涤后再加入HRP标记的二抗(稀释比例1∶10 000),37℃孵育1h。增强化学发光法
发光,胶片曝光,Image J图像分析软件分析Western blotting结果。以目标条带与β-actin条带灰度比值表示该蛋白的相对表达量。
1.2.5SOD活性和MDA检测:收集各干预条件下培养48 h的HUVECs,制成约10%的匀浆液,放置-20℃冰箱内,反复冻融3次或更多,直至显微镜下观察细胞已完全破碎,再以4 000 rpm离心15min,取上清液检测SOD活性和MDA浓度,操作严格按试剂盒说明书进行。
1.2.6流式细胞仪检测HUVECs凋亡:收集各干预条件下培养48h的HUVECs,通过流式细胞术进行凋亡检测。将培养液中悬浮的细胞和培养瓶内贴壁的细胞全部收集起来,离心后弃去上清,滴加200μl Annexin V-FITC结合液,震荡重悬细胞后再加入5μl Annexin V-FITC并混匀,在4℃下避光孵育15min。再在上述重悬细胞中加入5μl PI 染色液,混匀后于4 ℃下避光孵育5min,随后通过流式细胞仪(BD FACSCanto II, USA)检测。
1.3 统计学处理

2 结果
2.1 高糖对HUVECs氧化应激、自噬和凋亡的影响
研究结果显示,与对照组比较,高糖组HUVECs 的SOD活性下降,MDA水平显著升高(t=6.710、5.363, 均P<0.01),见表2。自噬相关基因检测结果表明,与对照组比较,高糖组HUVECs LC3II/LC3I比例增加,Beclin-1表达明显上升,而SQSTM1表达显著减少(t=5.023、6.486、15.070, 均P<0.01),见图1。流式细胞仪检测提示高糖组HUVECs的凋亡率显著增加(t=14.27,P<0.01),见图2。
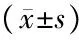
表2 各组HUVECs SOD活性和MDA水平比较
注:与对照组比较,1)P<0.01

注:与对照组相比,*P<0.05;#P<0.01

注:与对照组比较,*P<0.01
2.2 miR-17mimics转染对HUVECs miR-17表达的影响
各组HUVECs的miR-17表达水平差异有统计学意义(F=385.3,P<0.01)。与非转染组比较,miR-17 mimics组HUVECs的miR-17表达水平明显升高,较非转染组高66倍(t=19.630,P<0.01),而miR-control mimics组与非转染组miR-17表达无明显差异(t=1.732,P>0.05)。见图3。

注:与对照组比较,*P<0.01
2.3 miR-17过表达对高糖条件下HUVECs氧化应激、自噬和凋亡的影响
氧化应激指标和凋亡水平检测结果显示,在高糖条件下,miR-17mimics组HUVECs SOD活性较非转染组增加,MDA浓度较非转染组下降(t=6.831、7.797, 均P<0.01)。Western blotting检测结果表明高糖条件下,miR-17mimics组HUVECs的LC3II/LC3I比例及Beclin-1表达水平较非转染组上升,同时SQSTM1表达较非转染组减少(t=12.710、 5.155、11.740,均P<0.01)。流式细胞检测结果表明,高糖条件下miR-17mimics组HUVECs凋亡较非转染组减少(t=10.15,P<0.01)。此外,在高糖条件下,与miR-17 mimics组相比,miR-17 mimics+3-MA组HUVECs的SOD活性下降,MDA水平增加(t=8.115、6.910、7.783,均P<0.01),同时LC3II/I比例下降,Beclin-1表达减少,而SQSTM1水平增加(t=8.985、 17.980、13.060,均P<0.01),凋亡细胞比例增加(t=7.783,P<0.01)。见表3和图4、5。

表3 过表达miR-17对HUVECs SOD和MDA的影响
注: 与非转染组比较,1)P<0.01; 与miR-17 mimics组比较,2)P<0.01

注: 高糖条件下,与非转染组比较,*P<0.01;与miR-17 mimics组比较,#P<0.01

注:高糖条件下,与非转染组比较,*P<0.01;与miR-17 mimics组比较,#P<0.01
3 讨论
2型糖尿病(T2DM)是多种心血管疾病的危险因素,显著增加患者死亡率。内皮细胞功能失调是糖尿病血管病变的始动环节,而高血糖导致的活性氧簇形成增加是糖尿病引起内皮功能失调的主要因素[5],持续的氧化应激反应对内皮细胞产生损伤并导致其凋亡增加。氧化应激引起的内皮细胞损伤不仅直接导致血管结构改变,同时影响血管依赖于内皮调节的张力。临床研究表明,抗氧化剂可对糖尿病血管病变产生治疗作用。
本文结果显示,高糖组HUVECs的SOD活性下降,MDA水平升高,进一步证实高血糖可增加机体的氧化应激反应。自噬相关基因表达水平检测结果也表明高糖环境在一定程度上能激活自噬。HUVECs转染miR-17 mimics后,过表达miR-17可进一步增加高糖条件下HUVECs的自噬水平,抑制其氧化应激反应,减少凋亡;而HUVECs在经自噬抑制剂3-MA干预后,miR-17的上述作用显著减弱。这些结果表明上调内皮细胞自噬水平可减轻内皮细胞氧化应激反应,并具有抗凋亡作用。
miR-17参与多种细胞的自噬调节,结核分枝杆菌感染可通过下调miR-17的表达抑制巨噬细胞自噬,上调miR-17可靶向抑制Mcl-1和STAT3增强巨噬细胞自噬[6]。此外,在肝缺血再灌注损伤过程中,过表达miR-17可减少STAT3及其磷酸化水平,导致自噬上调,进而加重再灌注损伤[7]。这表明自噬的上调并不是都能产生保护作用,当组织或细
胞的损伤超过一定限度,自噬的上调反而会加速细胞凋亡。本文通过miR-17 mimics上调内皮细胞miR-17的表达,这种上调可实现对内皮细胞自噬的正向调节。
T2DM患者胰岛β细胞自噬水平增加[8]。这种自噬水平的上调可能是引起胰岛β细胞死亡的促进因素,而持续的血糖增高则可通过氧化应激等途径导致机体自噬增加。本研究结果显示,当内皮细胞处于高糖环境中时,其氧化应激反应水平增加,同时其自噬水平也有一定程度上调。当过表达miR-17时,高糖组的内皮细胞自噬水平被进一步上调,其凋亡有一定程度下降,但与正常组相比,内皮细胞的凋亡仍显著增加。这提示外源性因素引起的自噬上调不足以完全抵消非正常情况下的氧化应激反应引起的内皮细胞损伤。
综上,高糖条件下,在内皮细胞中过表达miR-17可通过增加其自噬水平减轻氧化应激反应,从而减少内皮细胞凋亡,具体的调节机制尚待进一步研究阐明。
◀
本文第一作者简介:
李罗成(1982年—),男,土家族,外科学博士,主治医师,研究方向为心血管疾病的发病机制
1 Gray SP, Di ME, Okabe J, et al. NADPH oxidase 1 plays a key role in diabetes mellitus-accelerated atherosclerosis.[J]. Circulation, 2013, 127(18):1 888-1 902.
2 Magenta A, Greco S, Gaetano C, et al. Oxidative stress and microRNAs in vascular diseases[J]. International journal of molecular sciences, 2013, 14(9):17 319-17 346.
3 Villard A, Marchand L, Thivolet C, et al. Diagnostic value of cell-free circulating microRNAs for obesity and type 2 diabetes: a meta-analysis.[J]. Journal of Molecular Biomarkers & Diagnosis, 2015, 6(6):251.
4 Dellago H, Bobbili MR, Grillari J. MicroRNA-17-5p: at the crossroads of cancer and aging-a mini-review[J]. Gerontology, 2017, 63(1):20.
5 Giacco F, Brownlee M. Oxidative stress and diabetic complications[J]. Circulation Research, 2010, 107(9):1 058-1 070.
6 Sasaki S, Inoguchi T. The Role of oxidative stress in the pathogenesis of diabetic vascular complications[J]. Diabetes & Metabolism Journal, 2012, 36(4):255-261.
7 Li S, Zhang J, Zhen W, et al. MiR-17 regulates autophagy to promote hepatic ischemia-reperfusion injury via suppression of Stat3 expression[J]. Liver Transplantation, 2016, 22(12):1 697-1 709.
8 Masini M, Bugliani M, Lupi R, et al. Autophagy in human type 2 diabetes pancreatic beta cells[J]. Diabetologia, 2009, 52(6):1 083-1 086.
猜你喜欢
航天电子对抗(2022年4期)2022-10-24
中国眼镜科技杂志(2019年9期)2019-11-11
天然产物研究与开发(2018年11期)2018-11-30
中国科技纵横(2018年2期)2018-11-29
中成药(2018年7期)2018-08-04
中成药(2018年6期)2018-07-11
中成药(2017年8期)2017-11-22
现代检验医学杂志(2016年4期)2016-11-15
中成药(2016年8期)2016-05-17
中国病理生理杂志(2015年8期)2015-12-21