
低温弱光下杭椒类辣椒不同世代间光合特性的比较
2018-03-03 03:26吕晓菡傅鸿妃吴根良陈建瑛
浙江农业科学 2018年2期
吕晓菡,傅鸿妃,吴根良,陈建瑛
(杭州市农业科学研究院 蔬菜研究所, 浙江 杭州 310024)
辣椒(CapsicumannuumL.)是茄科辣椒属,起源于中美洲和南美洲的热带与亚热带地区,为典型的喜温性蔬菜,不耐低温,是我国栽培面积最大的蔬菜种类之一[1-2]。杭椒类辣椒是杭州及周边地区传统优势辣椒品种,在全国负有盛名,一道“杭椒牛柳”已成为其金招牌。目前,杭椒类辣椒早就不再局限于江浙沪地区,在湖北、安徽、海南、福建、山东等多个省市及地区均有大规模种植。据统计,杭椒类辣椒品种种子每年销售已突破5 000 kg,种植面积达1万hm2以上,而且种植区域和面积还在不断扩大。杭椒类辣椒的最大优势在于品质好,种植效益高,特别是早春设施栽培,春节前后杭州市场辣椒的市场成交价超过10元·kg-1。然而,由于目前设施栽培大多以非加温的日光温室和塑料大棚为主,冬季和早春时节,江浙沪地区容易出现连阴雨天气。大量研究表明,当低温弱光逆境较轻时,植物叶片气孔关闭,光合作用减弱;当低温弱光逆境较严重时,光合作用碳同化关键酶RuBP羧化酶和其他一些酶的活性明显下降,并进一步减缓光合作用产物淀粉等的运输,造成对光合作用的反馈抑制,显著降低净光合速率,导致植株生长减慢,甚至停止生长[3-5]。因此,低温弱光胁迫一直是早春设施杭椒类辣椒生产中一种较为普遍的逆境,对其生产造成很大的限制。
光合作用是对低温弱光反应最敏感的生理过程之一。朱为民等[6]提出可将光合特性作为番茄设施专用品种选育的指标,王可玢等[7]提出叶绿素a荧光诱导动力学方法是鉴定番茄抗冷性的一个快速、灵敏和可靠的方法。近年来,有关低温弱光对辣椒光合速率的影响已有诸多报道,但对低温弱光下辣椒不同世代光合特性的差异却鲜有报道。本研究以杭椒类辣椒为材料,从植株光合特性入手,研究低温弱光下辣椒亲本与子代光合参数和叶绿素荧光参数的差异,以期初步获得探索杭椒类辣椒耐低温弱光遗传机理的新途径。
1 材料与方法
1.1 材料
选取杭椒类辣椒7份,分别为杭椒12号、杭椒2731、杭椒206、杭二♀-1-1-后-后、03-11-2-后-7-1-1、吉林2015、杭椒73-1-1。其亲缘关系如图1所示,其中,杭二♀-1-1-后-后、03-11-2-后-7-1-1为母本,吉林2015、杭椒73-1-1为父本,杭椒12号、杭椒2731、杭椒206为杂交F1代。供试辣椒种质于2016年8月中下旬在杭州市农业科学研究院蔬菜所试验基地内开始栽培。先将各辣椒种质进行穴盘育苗,然后移植入设施大棚内继续栽培备用。

图1 供试辣椒的亲缘关系
1.2 试验方法
低温处理:用温湿度仪每隔2 h测定棚内的温度,由于试验于2016年12月至翌年1月进行,叶绿素荧光参数测定前的15 d棚内昼夜平均温度始终低于或接近于5 ℃/3 ℃(昼温/夜温)。
弱光处理:在光合参数和叶绿素荧光参数测定前15 d,用遮阳网对大棚进行适当覆盖,并用光照测定仪测定光照强度,使棚内的光照强度始终在1 000 lx以下。
1.3 光合参数的测定
光合参数的测定参照文献[8]的方法。低温弱光处理10 d后,选取不同处理植株功能叶(从基部向上数第5叶或第6叶),利用便携式LI-6400光合仪(美国LI-COR公司),通过控光进行光响应曲线测定。每个材料每处理下测定重复3~4片叶,取平均值。在CO2浓度400 μmol·mol-1下,叶室光合有效辐射(PAR)设置梯度为0、20、50、100、200、500、1 000、1 500、2 000 μmol m-2·s-1,测定净光合速率(net photosynthesis rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)等参数。
1.4 叶绿素荧光参数的测定
叶绿素荧光参数的测定参照文献[9]的方法。低温处理10 d后,在不同低温弱光处理下,采用便携式LI-6400光合仪进行光强响应曲线、快速动力学曲线的测定,所有指标均重复3~5次,取平均值。选取不同处理植株功能叶(从基部向上数第5叶或第6叶)测定。测定前幼苗先暗适应1 h,然后关闭便携式LI-6400光合仪的内置光源,测定初始荧光(initial fluorescence,Fo)、最大荧光(maximal fluorescence,Fm)、PSⅡ最大光化学效率(Fv/Fm)。
1.5 数据分析
采用Microsoft Excel SAS统计软件,运用方差分析处理和分析数据,采用单因素方差分析(one-way ANOVA)比较不同处理之间的差异性,结果用平均值±标准差表示。
2 结果与分析
2.1 不同世代辣椒材料光合参数的差异
由表1可见,在相同的低温弱光逆境下,不论是不同世代间,还是亲本间、F1代间,辣椒植株的光补偿点(light compensation point,LCP)、光饱和点(light saturation point,LSP)、光饱和时光合速率等指标均出现明显差异。这可以初步说明,低温弱光胁迫可以通过直接影响光合机构的结构和活性来影响辣椒幼苗光合作用,在同一低温弱光逆境下,不同基因型的辣椒材料对逆境的耐受性有明显差异。另外,从表1中还可以发现,3个F1代的LCP、LSP、光饱和时光合速率指标大致居于父母本指标之间,其中,杭椒12号和杭椒206与其母本杭二♀-1-1-后-后、03-11-2-后-7-1-1更为接近。

表1 低温弱光下辣椒植株的光补偿点、光饱和点和光饱和时光合速率
注:测定时CO2浓度为(400 ± 10)μmol·mol-1,温度(10 ± 1)℃;表中数据为平均值±标准差。
逆境胁迫导致植物光合速率降低的原因包括气孔和非气孔因素。由图2可知,在低温弱光胁迫下,随着PAR的降低,叶片的Pn、Gs和Tr均减小,同时Ci逐渐增大,说明弱光下辣椒光合作用的主要限制因素是非气孔因素,如RuBP羧化酶受限制、无机磷限制、光化学机构受到破坏和光合产物运输减慢等原因减缓了光合作用,阻碍了光能对CO2的利用,造成细胞间隙CO2积累[10-11]。另外,对低温弱光下不同世代辣椒材料Pn、Ci、Gs、Tr变化进行分析发现,随着PAR的不断变化,3个F1代的Pn、Ci、Gs、Tr值始终居于其父母本之间或接近于其中一方(图2)。这说明,在低温弱光下,杭椒类辣椒对光的吸收能力可以在世代间稳定遗传。
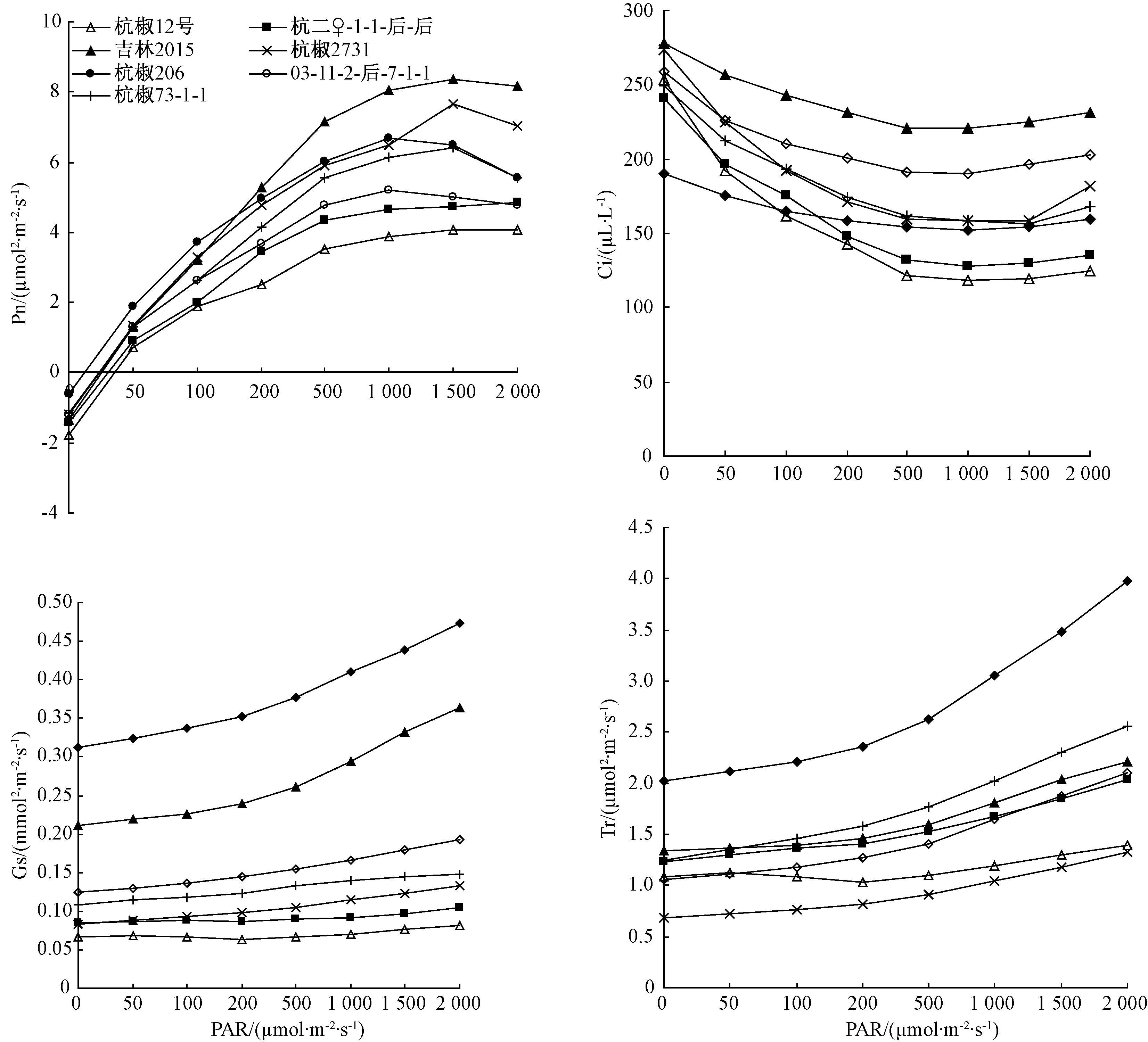
图2 不同辣椒幼苗在低温弱光下 Pn、Ci、Gs、Tr的变化
2.2 辣椒幼苗Fo、Fm、Fv/Fm的比较
基础荧光Fo表示PSⅡ反应中心全部开放,即原初电子受体Q全部氧化时的荧光水平,最大荧光Fm是PS反应中心完全关闭时的荧光产量,反映了通过PSⅡ的电子传递情况[12-13]。由图3可知,低温弱光下,3个F1代的Fo、Fm值都较接近于其亲本,推测在低温弱光下,杭椒类辣椒植株PSⅡ反应中心的电子传递能力可以在世代间稳定遗传。
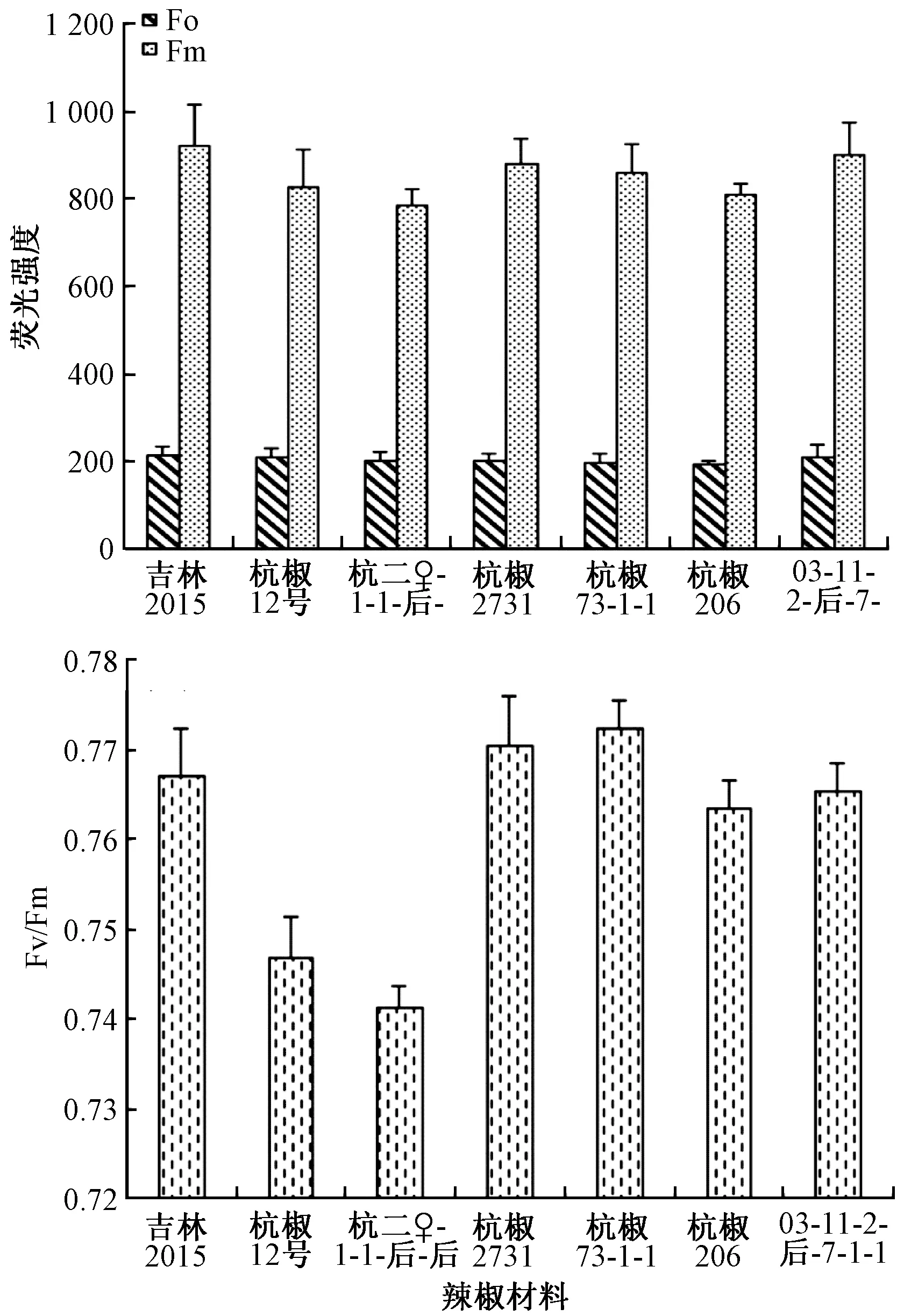
图3 不同辣椒幼苗在低温弱光下的Fo、Fm和Fv/Fm
最大光化学效率Fv/Fm指PSⅡ最大光化学效率(PSⅡ原初光能转化效率),它的变化代表PSⅡ光化学效率的变化,其值越低证明其发生光抑制的程度越高。由图3可知,3个F1代中,杭椒2731最大光化学效率最高,4个亲本材料中杭椒73-1-1的最大光化学效率最高,而杭椒73-1-1正是F1代杭椒2731的父本,这充分说明在低温弱光下,杭椒类辣椒植株的光能利用率和光合潜能可以在世代间稳定遗传。另外,母本同为杭二♀-1-1-后-后的F1代杭椒12号和杭椒2731的Fv/Fm值差异较大,而二者的父本(吉林2015、杭椒73-1-1)则差异较小。父本(杭椒73-1-1)相同的F1代,母本(杭二♀-1-1-后-后)Fv/Fm值低的F1代杭椒2731的Fv/Fm反而比母本(03-11-2-后-7-1-1)Fv/Fm值较高。说明F1代的光能利用率和光合潜能并不是简单地遗传自亲本的其中一方,而可能是父母本效应互作的结果。
3 讨论
低温弱光一直是制约越冬早春杭椒保护地生产的关键因子,因此,探索杭椒类辣椒耐低温弱光能力能否在世代间稳定遗传及其遗传机理,是选育耐低温弱光杭椒新品种的关键。光合参数和叶绿素荧光参数是鉴定植株耐低温弱光能力的重要指标。本研究发现,低温弱光胁迫导致杭椒类辣椒植株光合速率下降的因素是非气孔因素,可能是RuBP羧化受限制、光化学机构受到破坏等原因减缓了整个光合作用途径的运行,结果造成胞间CO2积累。另外,杭椒F1代植株的光合参数和叶绿素荧光参数均接近于其亲本,说明杭椒类辣椒植株在低温弱光下对光的吸收能力、电子传递能力及光能利用能力均可以被稳定地遗传给下一代,表明杭椒类辣椒
对低温弱光的耐受能力是可以在世代间稳定遗传的,亲本耐低温弱光能力强,通常其F1代也具有较高的耐低温弱光能力。另外,F1代的耐低温弱光能力并不是简单的遗传自亲本的其中一方,或者是父母本效应的简单叠加,而可能是一个复杂的父母本效应互作的结果,可能与父母本的特殊配合力有关,具体机理还需要继续研究。
[1] 徐伟慧, 王兰兰, 王志刚. 低温对辣椒幼苗生理生化特性的影响[J]. 甘肃农大学报, 2006, 41(4): 56-59.
[2] 王萍, 郭晓冬, 赵鹏. 低温弱光对辣椒叶片光合色素含量的影响[J]. 北方园艺, 2007(7): 15-17.
[3] 王永健, 张海英, 张峰, 等. 低温弱光对不同黄瓜品种幼苗光合作用的影响[J]. 园艺学报, 2001, 28(3): 230-234.
[4] 张红梅, 余纪柱, 金海军. 低温弱光对黄瓜植株生长、光合特性的影响[J]. 沈阳农业大学学报, 2006, 37(3): 339-342.
[5] 何勇, 符庆功, 朱祝军. 低温弱光对辣椒叶片光合作用、叶绿素荧光猝灭及光能分配的影响[J]. 核农学报, 2013, 27(4): 479-486.
[6] 朱为民, 朱龙英, 陆世钧, 等. 光合特性作为番茄设施专用品种选育指标的效应[J]. 上海农业学报, 2001, 17(4): 45-48.
[7] 王可玢, 赵福洪, 王孝宣, 等. 用体内叶绿素a荧光诱导动力学鉴定番茄的抗冷性[J]. 植物学报, 1996, 13(2): 29-33.
[8] 眭晓蕾, 张宝玺, 张振贤, 等. 不同品种辣椒幼苗光合特性及弱光耐受性的差异[J]. 园艺学报, 2005, 32(2): 222-227.
[9] 刘立云, 李艳, 杨伟波, 等. 不同品种油茶叶绿素荧光参数的比较研究[J]. 热带作物学报, 2012, 33(5): 886-889.
[10] ALLEN D J, ORT D R. Impacts of chilling temperatures on photosynthesis in warm-climate plants [J]. Trends in Plant Science, 2001, 6(1): 36-42.
[11] KRALL J P, EDWARDS G E, KRALL J P, et al. Environmental effects on the relationship between the quantum yields of carbon assimilation andinvivoPsⅡ electron transport in maize [J]. Functional Plant Biology, 1991, 18(3): 267-278.
[12] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学报, 1999, 16(4): 444-448.
[13] 孔海云, 张丽霞, 王日为. 低温与光照对茶树叶片叶绿素荧光参数的影响[J]. 茶叶, 2011, 37(2): 75-78.
猜你喜欢
江西通信科技(2022年2期)2022-08-08
福建农林大学学报(自然科学版)(2021年5期)2021-10-08
江苏农业科学(2019年2期)2019-08-13
天津农业科学(2019年1期)2019-07-23
农业科技与装备(2018年5期)2018-02-19
江苏农业科学(2017年1期)2017-02-27
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
福建农业科技(2016年10期)2016-03-07
少儿科学周刊·儿童版(2015年10期)2015-11-07
