
双孢蘑菇培养料隧道发酵过程中胞外降解酶活性的变化
2018-01-05 00:39:00朱燕华宋晓霞张津京李正鹏黄建春
上海农业学报 2017年6期
朱燕华,王 倩,宋晓霞,张津京,李正鹏,陈 辉,黄建春
(上海市农业科学院食用菌研究所,农业部南方食用菌资源利用重点实验室,国家食用菌工程技术研究中心,国家食用菌加工技术研发分中心,上海市农业遗传育种重点开放实验室,上海 201403)
双孢蘑菇培养料隧道发酵过程中胞外降解酶活性的变化
朱燕华,王 倩,宋晓霞,张津京,李正鹏,陈 辉,黄建春*
(上海市农业科学院食用菌研究所,农业部南方食用菌资源利用重点实验室,国家食用菌工程技术研究中心,国家食用菌加工技术研发分中心,上海市农业遗传育种重点开放实验室,上海 201403)
研究了双孢蘑菇培养料隧道式发酵各阶段主要降解酶活性的变化情况。结果表明:各降解酶的酶活性呈现不同的变化规律。纤维素降解酶分别在不同发酵阶段起作用,Cx酶活力在2次转仓时达到峰值,而C1酶与BG酶的酶活力分别在二次料与一次料中最高。半纤维素降解酶中的木聚糖酶活力呈先上升后下降的变化趋势,在2次转仓时酶活力最高,而木糖苷酶的活力一直较低。木质素降解酶(Lac、MnP与LiP)的活力变化趋势一致,均呈先上升后下降,再上升然后再下降的变化趋势,在2次转仓时各酶活力达到峰值。蛋白酶活力在1次转仓时最高,此后开始下降,在二次料中再次升高。淀粉酶活力仅在培养料开始发酵的时期较高,此后的酶活力一直较低。培养料中的木质素在1次转仓阶段降解最快,而纤维素和半纤维素降解最快的时期是在2次转仓阶段。
双孢蘑菇;培养料;发酵;酶活力
双孢蘑菇(Agaricus bisporus)是世界上栽培与消费最为广泛的食用菌[1]。栽培双孢蘑菇的原材料经过堆制后,形成有利于双孢蘑菇菌丝生长,而不利于其他杂菌生长的具有“选择性”的培养料。培养料的质量是影响双孢蘑菇产量与质量的关键因素[2],而堆肥中的微生物在培养料发酵过程中起着至关重要的作用[3]。双孢蘑菇堆肥过程中的微生物群落种类与数量不断演替,微生物活动时产生的胞外酶种类与活力大小也随之不断变化,促进堆肥各阶段培养料中的物质转化利用,从而形成了可促进双孢蘑菇菌丝生长的具有“选择性”的营养源[4]。目前对于不同双孢蘑菇栽培的培养料配方方面的研究较多[5],而对双孢蘑菇隧道式发酵过程中的微生物群落与功能、微生物的降解酶系变化规律方面的研究较少[6]。国内企业双孢蘑菇堆肥生产过程中仍主要凭借生产技术人员的经验来主观判断,导致目前我国的双孢蘑菇工厂化生产企业在堆肥的生产过程中,常出现培养料堆制过生或过熟、二次料氨气无法排尽抑制双孢蘑菇菌丝生长、堆肥的“选择性”差致使栽培过程中杂菌滋生,大面积减产等问题,引发重大的经济损失。本研究对工厂化隧道式发酵的双孢蘑菇培养料发酵过程中的主要物质降解酶活性进行测定,分析发酵各阶段的物质降解转化特点,以期为进一步研究微生物群落在双孢蘑菇培养料发酵中的功能及合理制定双孢蘑菇堆肥技术提供参考。
1 材料与方法
1.1 供试菌株
双孢蘑菇(A.bisporus)A15菌种,由美国Sylvan公司提供。
1.2 试验材料
堆肥配方为:稻草 23 t、麦草 15 t、棉籽壳 2 t、菜籽饼 2 t、石膏 2.5 t、碳酸钙 1 t。
1.3 培养料发酵
试验地点为上海市农业科学院金山双孢蘑菇种植基地。双孢蘑菇的培养料发酵采用二次发酵工艺。全麦草配方中的麦草预湿后与鸡粪、石膏等均匀混合后,在一次发酵槽中进行发酵。60%稻草配方中的麦草提前2 d进行预湿,然后再与稻草、鸡粪、石膏、棉籽饼等均匀混合。此后每隔2—3 d转仓一次,转仓3—4次后,完成一次发酵,持续时间约14 d左右。将一次料运至二次发酵隧道中,进行密闭式发酵,通过调节通风量控制培养料的温度,依次经过均温、巴氏消毒、空气调节及冷却阶段,测定氨气浓度低于0.005‰时,即可降温完成二次发酵过程,持续时间约7 d左右。二次料运至菇房中,播撒双孢蘑菇菌种,播种量为培养料干重的0.7%,采用上料机将培养料放于床架上进行发菌、出菇等栽培管理。
1.4 取样方法
取样阶段为:进仓、1次转仓、2次转仓、3次转仓、4次转仓、一次料、二次料共7个取样时期。
取样时,将发酵隧道中的样品分为3段取样,即隧道前中后3个位置。每个位置间隔距离约5 m。在每个取样位置的剖面,按高度分为上(顶部往下10—20 cm)、中、下(地面以上10—20 cm)3个部分,五点梅花形取样,每个点取样100 g,将每个剖面的5个样品混匀后,采用四分法取样,每个取样时期的培养料样品设3个重复。
1.5 测定方法
粗酶液提取:参照杨新美[7]的方法,从料面下2—3 cm处均匀取样5 g,加入25 mL水,25℃摇床浸取1 h,转速为150 r/min,然后4 000 r/min、5℃离心10 min,取上清液(或用四层纱布过滤,过滤液即为粗酶液),立即测定或于-30℃冰箱中保存待测。
纤维素酶活性测定:纤维素酶是一种复合酶系,由外切葡聚糖纤维二糖水解酶(C1酶)、内切葡聚糖酶(Cx酶)和β-葡聚糖苷酶(BG)组成,Cx酶、BG酶、C1酶活性的测定采用DNS法[8]。Cx酶与BG酶活力单位为1 mL酶液1 h产生1μmol葡萄糖为1个酶活力单位(U),C1酶活力单位为1 mL酶液24 h产生1μmol葡萄糖为1个酶活力单位(U)。
半纤维素酶活性测定:半纤维素降解酶主要为内切木聚糖酶与β-木糖苷酶。木聚糖酶与β-木糖苷酶活性的测定分别参照赵超等[8]与Cai等[9]。半纤维素的酶活力单位为1 mL酶液1 h产生1μmol葡萄糖为 1个酶活力单位(U)[8]。
木质素酶活性测定:木质素降解酶主要包含3个酶系:漆酶(Lac)、锰过氧化物酶(MnP)和木质素过氧化物酶(LiP),木质素降解酶活性的测定方法参照 Buswell等[10]。Lac酶活力单位(U)=(A×106)/(∑420×d),其中 A=60 s内吸收值的增加量,d=光路(cm),比色皿的直径;消光系数∑420=3.6×104/(mol·cm)[11]。MnP酶活力单位(U)=(A×106)/(∑240×d),其中 A=60 s内吸收值的增加量,d=光路(cm);消光系数∑240=8.1×103/(mol·cm)[12]。LiP酶活力单位(U)为1 min内在310 nm处引起吸光度值变化0.1所需的酶量[13]。
蛋白酶活力测定:参照倪新江等[14]的方法,蛋白酶活力单位(U)为每克干培养物30 min内改变0.1个光密度值。淀粉酶活力测定:参照王玉万等[15],淀粉酶活力单位(U)为每克干物质30 min内反应生产1 mg葡萄糖为1个酶活力单位。
2 结果与分析
2.1 培养料发酵过程中纤维素酶活性的变化
Cx酶作用于纤维素分子内部的非结晶区,随机水解β-1,4糖苷键,产生大量有非还原端的小分子纤维素。在培养料发酵过程中,堆肥中的Cx酶活力呈先上升后下降的变化趋势,在2次转仓时酶活力达到高峰,为12.4 U/g,此后开始下降。C1酶主要作用于纤维素线状分子的末端,水解β-1,4糖苷键,每次切下一个纤维二糖分子。C1酶活力在一次发酵过程中普遍较低,在二次料中的C1酶活力达到峰值,为8.24 U/g;BG酶的作用主要为将纤维二糖水解成葡萄糖分子,BG酶活力呈先上升后下降的趋势,在一次发酵结束时达到峰值,二次料发酵结束后的BG酶活力再次降低。上述结果表明,纤维素降解复合酶系中的各种酶在双孢蘑菇隧道式发酵不同阶段发挥降解功能。
2.2 培养料发酵过程中半纤维素酶活性的变化
在双孢蘑菇堆肥发酵过程中,降解半纤维素的主要酶为内切木聚糖酶,其作用主要为随机切断木聚糖骨架,产生木寡糖,降低聚合度。木聚糖酶活力呈先下降后上升,此后再下降的趋势,木聚糖酶的峰值出现在2次转仓开始时,酶活力达162.9 U/g,在3次转仓时迅速下降至53.5 U/g。β-木糖苷酶的作用是将木寡糖和木二糖分解为木糖。相对于木聚糖酶,在双孢蘑菇培养料发酵过程中的β-木糖苷酶活力较低,表明在双孢蘑菇培养料发酵过程中,木聚糖酶在半纤维素降解中起主要作用。

图1 培养料发酵过程中纤维素酶活性的变化Fig.1 Changes of cellulase activity during com post fermentation
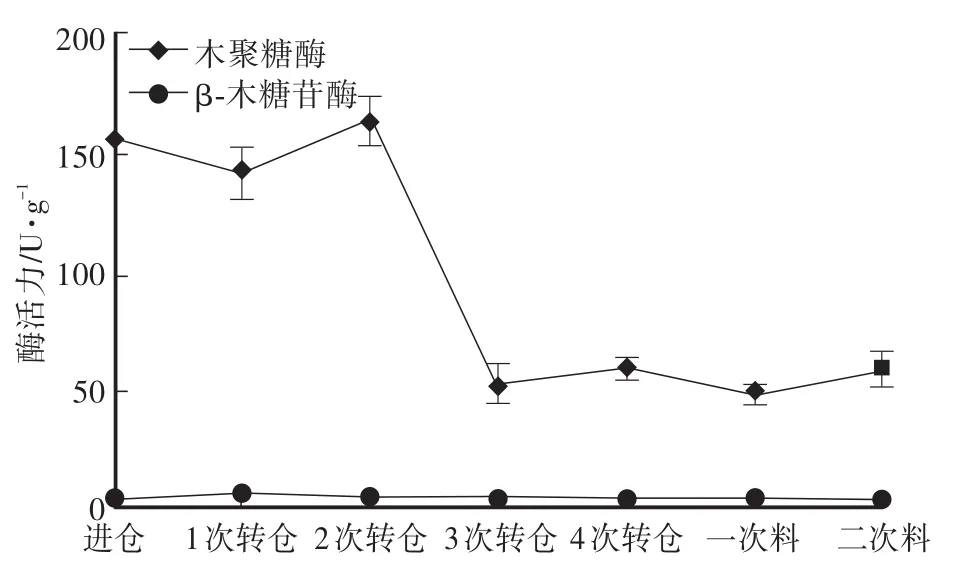
图2 培养料发酵过程中半纤维素酶活性的变化Fig.2 Changes of hem icellulose activity during compost fermentation
2.3 培养料发酵过程中木质素酶活性的变化
培养料发酵过程中的木质素降解复合酶系呈类似的变化趋势,Lac、MnP与LiP这3种酶的活力在刚开始发酵时均逐渐上升,在第2次转仓开始时达到峰值,此时酶活力最高的为Lac,其次为MnP,LiP的酶活力最低。第3次转仓时,这3种木质素降解酶的活力均开始下降,在一次发酵结束时这3种酶的活力再次升高,二次发酵结束后,酶活力均再次下降。
2.4 培养料发酵过程中蛋白酶与淀粉酶活性的变化
双孢蘑菇培养料中富含蛋白质,在发酵过程中蛋白酶活力也呈规律性变化,总体呈先上升后下降再上升的趋势。蛋白酶活力的峰值出现在1次转仓时,酶活力为17.5 U/g,表明此阶段微生物大量转化利用培养料中的氨基酸等小分子。此后,蛋白酶活力开始迅速下降,在一次发酵结束时最低,为0.6 U/g,但二次发酵结束时,堆肥中的蛋白酶活力又再次升高,达10.7 U/g,表明二次料中的微生物对蛋白质的降解利用较为旺盛。在进一次发酵仓时,培养料中的淀粉酶活力最高,此后逐渐下降,在2次转仓时降至最低,酶活力为0.36 U/g,此后堆肥各阶段的淀粉酶活力一直维持在较低的水平,表明在刚开始发酵时,微生物将易被利用的淀粉作为碳源,培养料中的淀粉等物质在2次转仓前就已基本被微生物所利用消耗。

图3 培养料发酵过程中木质素酶活性的变化Fig.3 Changes of ligninase activity during compost fermentation

图4 培养料发酵过程中蛋白酶与淀粉酶活性的变化Fig.4 Changes of protease activity and amylase activity during compost fermentation
2.5 培养料发酵各阶段培养料的木质纤维素降解率
双孢蘑菇培养料中的木质素在二次发酵结束时的降解率为27.0%,其中在1次转仓阶段降解速率最快,木质素降解率约占整个发酵阶段降解率的42.2%。培养料中纤维素与半纤维素在二次发酵结束时的降解率分别为57.9%与59.8%,其中在2次转仓阶段的降解速率最快,降解率分别为38.4%与39.0%。
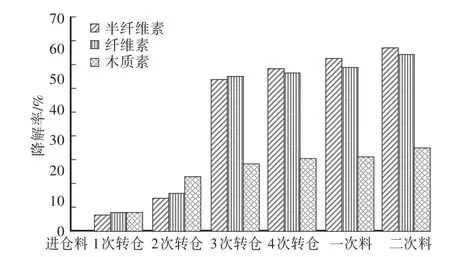
图5 培养料发酵过程中木质纤维素降解率的变化Fig.5 Changes of lignocellulose degradation rate during com post fermentation
3 讨论
本研究表明,该批次的培养料栽培双孢蘑菇的三潮菇总产量为25.4 kg/m2,表明培养料的质量较好。在双孢蘑菇培养料隧道式发酵过程中,培养料中主要物质降解酶系的酶活力呈现不同的变化规律,在2次转仓开始时的木质素降解酶(Lac、MnP与LiP)、纤维素降解酶(Cx酶)及半纤维素降解酶(木聚糖酶)的酶活力均达到峰值,表明在堆肥过程中2次转仓开始时的微生物对培养料中主要成分的降解转化能力最强。此外,一次发酵料中的3种木质素降解酶与BG酶活力也较高。二次发酵料中可能由于存在大量的细菌残体蛋白[16],有利于其他微生物进行生长利用,因而二次料的蛋白酶活力也较高。李晓博等[6]采用传统方法进行双孢蘑菇培养料发酵过程中酶活性变化的研究表明,半纤维素酶活力在第2次翻堆时最高,纤维素酶活力在第4次翻堆时最高,而漆酶活力在第3次翻堆时最高,酶活力峰值的出现时间均低于本研究中利用隧道进行培养料发酵,这可能是由于隧道式发酵环境更有利于微生物群落快速繁殖与演替,从而加速了发酵进程。侯晓伟等[17]研究表明,利用大麦秸秆作为原材料进行双孢蘑菇培养料发酵,培养料中的纤维素和半纤维素降解最快的时期在第2次翻堆后,这与本研究结果一致。在双孢蘑菇培养料发酵过程中,纤维素降解酶系中的各降解酶在培养料发酵过程中呈现不同的变化规律,各自在特定发酵阶段发挥降解作用,培养料中纤维素降解最快的时期Cx酶活力也处于峰值,表明Cx酶可能在纤维素的降解中起主导作用。木聚糖酶是培养料中半纤维素降解的主要酶。木质素降解酶系中的各种酶变化规律一致,这3种酶在木质素降解过程中存在较好的协同关系,共同作用从而完成各阶段的木质素降解。根据本研究结果,结合各种降解酶活性的变化规律,在双孢蘑菇培养料隧道式发酵过程中应使培养料温度尽快升高,从而使木质纤维素的相关降解酶活性在较短时间内达到峰值,这对于缩短培养料发酵时间,减少培养料中的物质能量损耗具有重要作用。培养料发酵各阶段酶活性变化特性的研究也为进一步了解双孢蘑菇培养料隧道式发酵中微生物对物质的降解转化规律及提高双孢蘑菇培养料发酵的质量等提供参考。
[1]BERENDSEN R,KALKHOCE S,LUGONES L,et al.Effects of themushroom-volatile 1-octen-3-ol on dry bubble disease[J].Appl Microbiol Biotechnol,2013,97(12):5535-5543.
[2]KARIAGA M.Important factors in composting for production of high yields in button mushrooms and Agaricus bitorquis(Quel)Saccardo[J].African Crop Science Conference Proceedings,2005,7:1273-1277.
[3]郭亚萍,张国庆,陈青君,等.双孢蘑菇堆肥过程中细菌群落结构分析[J].应用与环境生物学报,2014,20(5):832-839.
[4]LIYAMA K,STONE B,Macauley B J.Compositional changes in compost during composting and growth of Agaricus bisporus[J].Appl Environ Microbiol,1994,60(5):1538-1546.
[5]陈茜,王建立,郭亚萍,等.不同培养料配方对双孢蘑菇产量的影响[J].中国农学通报,2014,30(4):185-189.
[6]李晓博,李晓,李玉.双孢蘑菇生产中木质素、纤维素和半纤维素的降解及利用研究[J].食用菌,2009(2):6-9.
[7]杨新美.食用菌研究法[M].北京:中国农业出版社,1998.
[8]赵超,高兆银,刘美珍.不同配方蔗渣培养基对平菇纤维素酶活性和产量的影响[J].浙江农业科学,2010(1):31-34.
[9]CAIY,BUSWELL JA,CHANG S.Cellulases and hemicellulases of Voluarielka volvacea and the effect of Tween 80 on enzyme production[J].Mycological Research,1994,98:1019-1024.
[10]BUSWELL J,CAIY,CHANG S,et al.Lignocellulolytic enzyme profiles of ediblemushroom fungi[J].World JMicrobiol Biotechnol,1996,12(5):537-542.
[11]ISIKHUEMHEN O,MIKIASHVILIN,KELKAR V.Application of solid waste from anaerobic digestion of poultry litter in Agrocybe aegerita cultivation:mushroom production,lignocellulolytic enzymes activity and substrate utilization[J].Biodegradation,2009,20:351-361.
[12]AITKEN M,IRVINE R.Characterization of reactions catalyzed by manganese peroxidase from Phanerochaefe chysosporium[J].Archives of Biochemistry and Biophysics,1990,276:405-414.
[13]ARORA D,Gill R.Comparison of two assay procedures for lignin peroxidase[J].Enzyme and Microbal Technology,2001,28:602-605.
[14]倪新江,丁立孝,潘迎捷,等.姬松茸在两种培养基上生长期间九种胞外酶活性变化[J].菌物系统,2001,20(2):222-227.
[15]王玉万,王云.构菌栽培过程中对木质纤维素的降解的和几种多糖分解酶活性的变化[J].微生物学通报,1989,16(3):137-140.
[16]SMITH J.Factorsaffecting the selectivity of composts suitable for the cultivation of Agaricus species[D].England:University of London,1994.
[17]侯晓伟,王晓巍,陈年来,等.双孢蘑菇培养料发酵微生物变化对其理化性质的影响研究[J].中国农学通报,2014,30(25):111-115.
Changes of extracellular degrading enzyme activity during the tunnel fermentation of Agaricus bisporus com post
ZHU Yan-hua,WANG Qian,SONG Xiao-xia,ZHANG Jin-jing,LIZheng-peng,CHEN Hui,HUANG Jian-chun*
(Institute of Edible Fungi,Shanghai Academy of Agricultural Sciences;Key Laboratory of Edible Fungi Resources and Utilization(South),Ministry of Agriculture,P.R.China;National Engineering Research Center of Edible Fungi;National R&D Center for Edible Fungi Processing;Key Laboratory of Agricultural Genetics and Breeding of Shanghai,Shanghai201403,China)
The changes ofmain degrading enzyme activities in different stages of tunnel culture of Agaricus bisporus compost were studied.The results showed that the activities of different degrading enzymes showed different change rules.Cellulose degrading enzymes acted at different fermentation stages,and Cx enzyme activity reached the peak at the second turn,while the activities of C1 and BG enzymeswere the highest in the secondary materials and primarymaterials respectively.Hemicellulose degrading enzymes include xylanase and xylosidase.Xylanase activity increased firstand then decreased,and reached the highestat the second turn,while the activity of xylosidase was always low.The change trend of lignin degrading enzymes(Lac,MnP and LiP)were the same,and they all increased first,then decreased,then increased,and then decreased,and reached the peak at the second turn.Protease activity was the highest at the first turn,then decreased,and increased again in the secondarymaterials.Amylase activity was only higher at the beginning of fermentation,and then the enzyme activity was always low.Lignin in the culture medium degraded fastest at the first turn,while cellulose and hemicellulose degraded fastest at the second turn.
Agaricus bisporus;Culturematerial;Fermentation;Enzyme activity
2016-04-11
农业部公益性(农业)行业科研专项(201503137);上海市科技兴农推广项目 [沪农科推字(2014)第2-2号];上海市科学技术委员会科研计划项目(15391900200)
朱燕华(1982—),女,博士,助理研究员,主要从事双孢蘑菇栽培与生理研究。E-mail:zhuyanhua@saas.sh.cn,Tel:021-62200538
*通信作者,E-mail:jianmushroom@163.com
S646
A
1000-3924(2017)06-006-05
闫其涛)
猜你喜欢
林产化学与工业(2022年4期)2022-09-13 09:47:12
中国食用菌(2021年10期)2021-11-04 06:23:26
农家之友(2020年5期)2020-06-28 02:16:46
湖南饲料(2019年5期)2019-10-15 08:59:10
食用菌(2018年5期)2019-01-08 02:52:34
现代食品(2018年4期)2018-02-18 15:50:01
科学种养(2017年8期)2017-08-10 20:46:40
食用菌(2017年3期)2017-05-24 06:52:18
西藏科技(2016年8期)2016-09-26 09:00:59
中国酿造(2016年12期)2016-03-01 03:08:21
