
山东省动物源沙门氏菌MLST和血清分型与分布研究
2017-10-09 05:04郭树源李璐璐刘玉洁王永明常维山刘玉庆
中国人兽共患病学报 2017年9期
赵 翠,张 庆,郭树源,李璐璐,刘玉洁,王永明,常维山,刘玉庆
DOI:10.3969/j.issn.1002-2694.2017.09.008
山东省动物源沙门氏菌MLST和血清分型与分布研究
赵 翠1,张 庆2,郭树源3,李璐璐2,刘玉洁4,王永明5,常维山3,刘玉庆2
目的比较动物源沙门氏菌多位点序列分型(MLST)与血清分型的差别,获得动物源沙门氏菌在山东省的分布规律。方法从山东部分地区分离78株鸡源沙门氏菌、56株鸭源沙门氏菌和20株猪源的沙门氏菌,PCR扩增七段沙门氏菌内部保守的片段序列进行多位点序列分型,同时用玻片凝集法分析其血清型。结果血清分析结果表明,鸡源沙门氏菌检出6种血清型,其中肠炎沙门氏菌占88.5%、印第安纳沙门氏菌占5.1%、汤卜逊沙门氏菌占2.6%、鼠伤寒沙门氏菌占1.3%、山夫登堡沙门氏菌占1.3%、阿格玛沙门氏菌占1.3%;鸭源沙门氏菌检出2种血清型,其中肠炎沙门氏菌占67.9%、鼠伤寒沙门氏菌占32.1%;猪源沙门氏菌检出3种血清型,其中鼠伤寒沙门氏菌占65%、德贝儿沙门氏菌占20%、肠炎沙门氏菌占15%。通过MLST分型,鸡源检出7种ST型:ST11、ST19、ST26、ST128、ST14、ST17和New1;鸭源检出3种ST型:ST11、ST19和New2;猪源检出3种:ST34、ST40和ST3007。结论整体来看,分离菌的血清型和ST型数量比较接近,二者分型能力相近。
沙门氏菌;血清型;MLST;分型
沙门氏菌(Salmonella)是肠杆菌科中的一个大属,是具有重要公共卫生学意义的人兽共患病原菌,全世界已发现2500多个血清型[1],中国已发现292个血清型[2]。近年来,该菌在畜禽中的带菌和发病状况,以及畜产品中毒的发生规律在一些发达国家受到高度重视,已经有详细的调查监测报告。我国畜禽养殖规模扩大和感染的病例持续上升,也应当引起养殖场和公共卫生的共同重视[3]。
沙门氏菌类别众多且分类方法也很多,伴随着沙门氏菌的传播逐渐由暴发开始转向多点散发,仅仅依靠血清学分型的方法已经不能完成病原体的溯源工作,而多位点序列分型(Multilocus sequence typing, MLST)是近几年通行的一种分子生物学分型方法[4-6]。它的分型原理是选择某一种菌的几段管家基因作为分型的目标基因,分别设计引物扩增并测序,根据菌株每个基因的序列信息分配等位基因序号,把该菌株所有基因的等位基因序号合并在一起组成一个等位基因谱,并给这个等位基因谱分配一个唯一的编号作为该分离株的序列型别[7]。该方法克服了PFGE和flaA难以分型及溯源的缺点,具有分辨率高、数据可靠、重复性好、结果便于不同实验室进行比较,从而使全球的分子流行病学数据标准化等优点[8-9]。
当前,有关山东省沙门氏菌多位点序列分型的数据资料比较少,因此本实验从山东省不同地区的养殖场收集鸡源、鸭源和猪源三种不同动物源沙门氏菌进行分离鉴定,并选取沙门氏菌7对保守管家基因aroC、dnaN、hemD、hisD、purE、sucA、thrA,对沙门氏菌进行多位点基因序列分型,初步建立山东地区沙门氏菌MLST分型数据库。同时检测了分离菌株的血清型,并比较两种方法的特点,从而分析山东地区的沙门氏菌的溯源和流行情况,为沙门氏菌病的防控提供参考。
1 材料与方法
1.1 样品来源 从山东省不同地区养殖场不同动物源收集动物的肛拭子、脑、心、肝、粪水和鸡胚样品,样品来源如表1。取样时每个养殖场舍取5份样品分析。
表1 样品来源
Tab.1 Samples sources
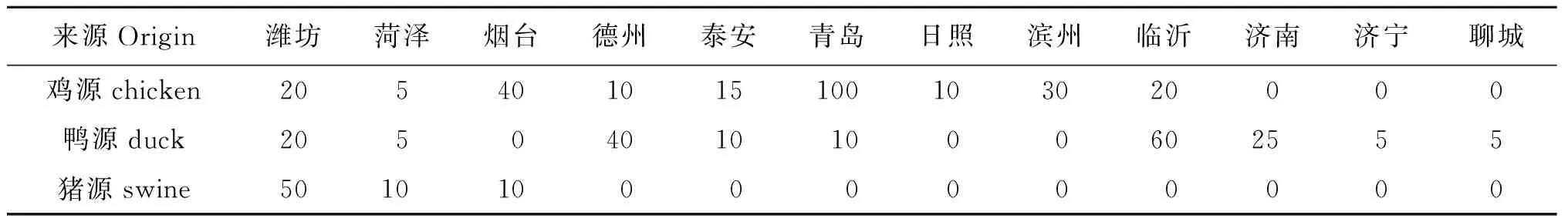
来源Origin潍坊菏泽烟台德州泰安青岛日照滨州临沂济南济宁聊城鸡源chicken205401015100103020000鸭源duck205040101000602555猪源swine501010000000000
1.2 细菌的分离鉴定和保存 将得到的样品进行增菌培养,然后用无菌的接种环蘸取增菌液,划线接种于沙门氏菌的科玛嘉显色培养基,于37 ℃培养 12~24 h。挑取疑似菌落,在SS培养基和麦康凯培养基上进行第二次划线分离,37 ℃温箱中培养24 h,进行再次鉴定,然后在LB固体培养基上划线培养单菌落,接种LB液体培养基,37 ℃摇床过夜培养。吸取700 μL培养的菌液加入到300 μL的灭菌的无菌甘油中振荡混匀,放入-20 ℃冷冻保存备用。
1.3 沙门氏菌血清分型的鉴定与分析 将沙门氏菌接种到固体琼脂和软琼脂平板上,37 ℃过夜培养。参照沙门氏菌检验标准GB/T 4789.4-2010,利用沙门氏菌属诊断血清试剂盒(宁波天润,批号20170101)进行试验。
挑取固体琼脂平板上的菌落进行O相血清凝集:先在洁净的玻片上滴加1~2滴诊断血清,然后用洁净的枪头挑取固体琼脂平板上的菌落与滴加的血清混匀,轻轻摇动玻片,1 min之内肉眼判断结果是否为O抗原型。如果O抗原不凝集,将菌株接种在琼脂含量较高的(如2%~3%)培养基上再检查,如果是由于Vi抗原的存在而阻止了O抗原凝集反应时,可挑取菌落在1 mL生理盐水中做成浓菌液,在酒精灯火焰上煮沸后检查。之后再挑取软琼脂周边扩散菌苔进行H相血清诊断,确定鞭毛抗原。
根据O相与H相的凝集结果,查阅沙门氏菌的血清分型表,确定沙门氏菌的抗原式和血清型。
1.4 沙门氏菌多位点序列分型(MLST)
1.4.1 MLST扩增引物的设计 根据PubMLST数据库以及NCBI上公布的基因片段设计7个管家基因hisD、purE、sucA、aroC、thrA、dnaN、hemD的引物序列,由上海生工生物工程有限公司合成。引物序列如下表2所示:
表2 沙门氏菌7个管家基因的引物序列
Tab.2 Primer sequences of Salmonella seven housekeeping genes

基因Gene引物Primer基因大小LengthofgenethrAF5'-GTCACGGTGATCGATC-CGGT-3'852bpR5'-CACGATATTGATATT-AGCCCG-3'aroCF5'-CCTGGCACCTCGCGC-TATAC-3'826bpR5'-CCACACACGGATCGTG-GCG-3'dnaNF5'-ATGAAATTTACCGTT-GAACGTGA-3'833bpR5'-AATTTCTCATTCGAGAG-GATTGC-3'sucAF5'-AGCAC-CGAAGAGAAACGCTG-3'643bpR5'-GGTTGTTGATAACGAT-ACGTAC-3'hisDF5'-TCGCGTCTGTCGGTCTG-TAT-3'755bpR5'-GGCGCAGTATAGCCAT-AGGT-3'hemDF5'-ATGAGTATTCTGATCAC-CCG-3'666bpR5'-ATCAGCGACCTTA-ATATCTTGCCA-3'purEF5'-GACACCTCAAAAG-CAGCG-3'510bpR5'-CGAGAACGCAAACTT-GCTTC-3'
1.4.2 DNA的提取和PCR扩增 用细菌基因组DNA提取试剂盒(购自北京天根生物科技有限公司),提取细菌DNA。多位点序列分型的PCR的反应体系:10×Buffer(Mg2+)2.5 μL,dNTPs (2.5 mmol/L) 2 μL,上游引物(25 mnol/L) 0.5 μL,下游引物(25 mnol/L) 0.5μL,模板DNA 1 μL,EasyTaq DNA polymerse 0.5 μL,ddH2O 18 μL,总共25 μL的体系。PCR 扩增程序:94 ℃预变性 10 min,94 ℃变性50 s,55 ℃退火50 s,72 ℃延伸50 s,32个循环,72 ℃延伸10 min,4 ℃保存。
1.4.3 测序及测序结果分析 PCR结束之后进行凝胶电泳,并将胶回收产物交上海生物工程技术有限公司进行双向测序。
将测序结果用DNA Star软件根据Pub MLST的相关要求进行修正,使7个管家基因均符合国际上关于沙门氏菌多位点序列分型的序列大小要求,将修正好的7个管家基因序列与MLST数据库中序列进行比对分析,分别获取七个管家基因位点的等位基因数值,并形成相应的等位基因谱,判断其序列型。
2 结果与分析
2.1 分离菌株 总共分离鉴定得到154株沙门氏菌,动物来源和地理分布见表3。
从分离结果来看,不同地区的分离率有差异,这应该与当地的养殖环境有关。另外,个别地区分离得到的菌株较少,这是由于取样数量较少造成的。
2.2 血清型分型结果 154株动物源沙门氏菌中肠炎沙门氏菌数量最多,共有110株,占总的菌株数71.4%(110/154);其次是鼠伤寒沙门氏菌32株、印第安纳沙门氏菌4株、德贝儿沙门氏菌4株、汤卜逊沙门氏菌2株、山夫登堡沙门氏菌1株、阿格玛沙门氏菌1株。总体来看肠炎型的沙门氏菌占据多数,其次为鼠伤寒沙门氏菌,其他的血清型呈现零星分布。具体分布如表4。
鸡源总共检测出6种血清型,其中肠炎沙门氏菌共有68株,占菌株的88.5%(68/78);其次印第安纳沙门氏菌4株、汤卜逊沙门氏菌2株、鼠伤寒沙门氏菌1株、山夫登堡沙门氏菌1株、阿格玛沙门氏菌1株。
鸭源只检测出2种血清型:肠炎沙门氏菌38株,占菌株的67.9%(38/56);鼠伤寒沙门氏菌18株,占菌株的32.1%(18/56)。鼠伤寒沙门氏菌比例明显高于鸡源沙门氏菌。
猪源总共检测出3种血清型,其中鼠伤寒沙门氏菌的共有13株,占能分型菌株的65%(13/20);德贝儿沙门氏菌4株、肠炎沙门氏菌3株。
猪源鼠伤寒沙门氏菌比例最多,而肠炎沙门氏菌的数量则较少,这与鸡源和鸭源分离情况不同。针对不同来源的血清型进行统计分析结果显示存在统计学差异性(P=0.048)。
表3 各地区分离情况统计
Tab.3 Strains separated from different regions
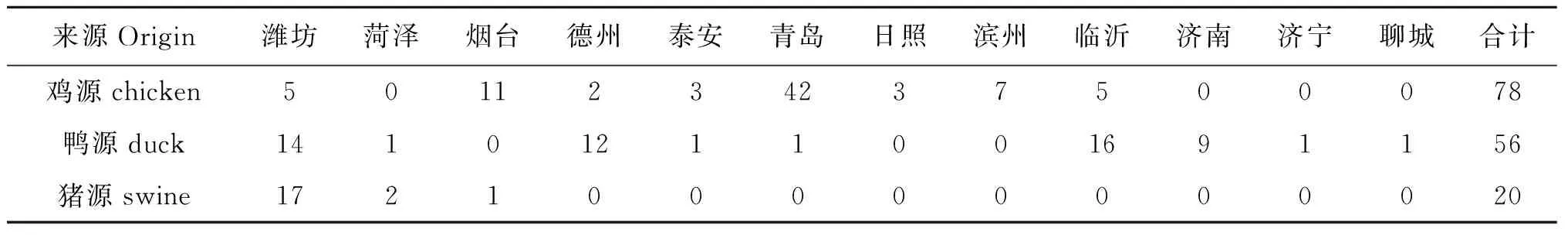
来源Origin潍坊菏泽烟台德州泰安青岛日照滨州临沂济南济宁聊城合计鸡源chicken5011234237500078鸭源duck14101211001691156猪源swine172100000000020
表4 不同动物源沙门氏菌血清型鉴定结果
Tab.4 Serotype identification result of different animal origin Salmonella
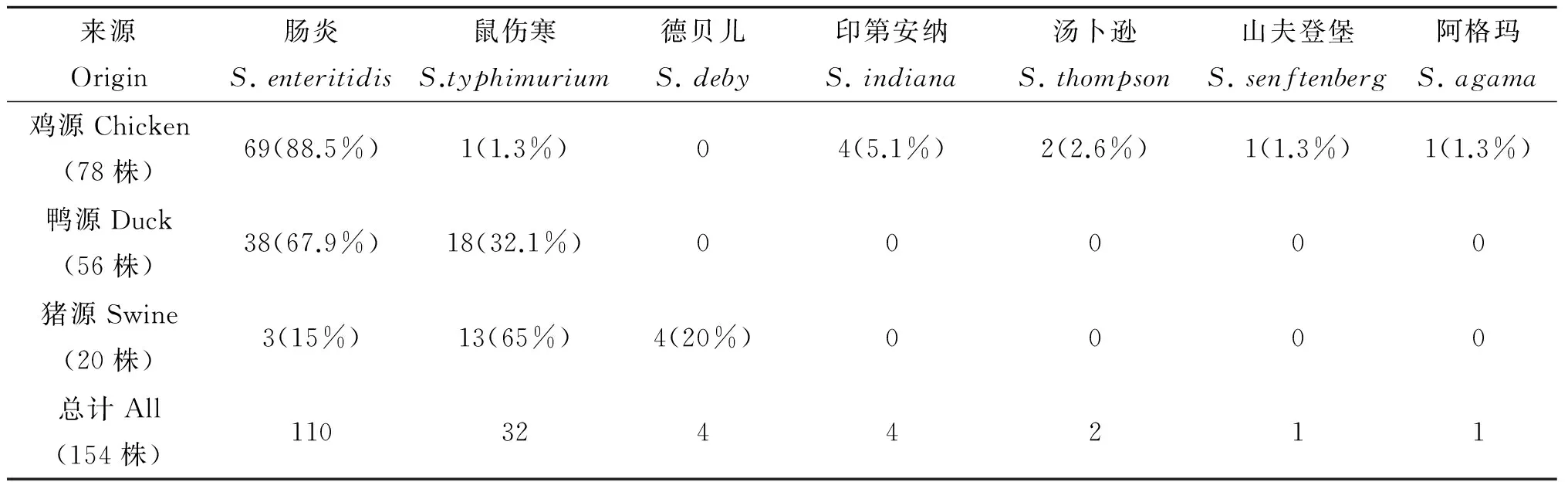
来源Origin肠炎S.enteritidis鼠伤寒S.typhimurium德贝儿S.deby印第安纳S.indiana汤卜逊S.thompson山夫登堡S.senftenberg阿格玛S.agama鸡源Chicken(78株)69(88.5%)1(1.3%)04(5.1%)2(2.6%)1(1.3%)1(1.3%)鸭源Duck(56株)38(67.9%)18(32.1%)00000猪源Swine(20株)3(15%)13(65%)4(20%)0000总计All(154株)1103244211
2.3 多位点序列分型结果 利用凝胶成像系统观察,可见沙门氏菌的7个管家基因都能扩增到相应的目的片段(如图1所示)。目的的片段大小符合表1的预期值。
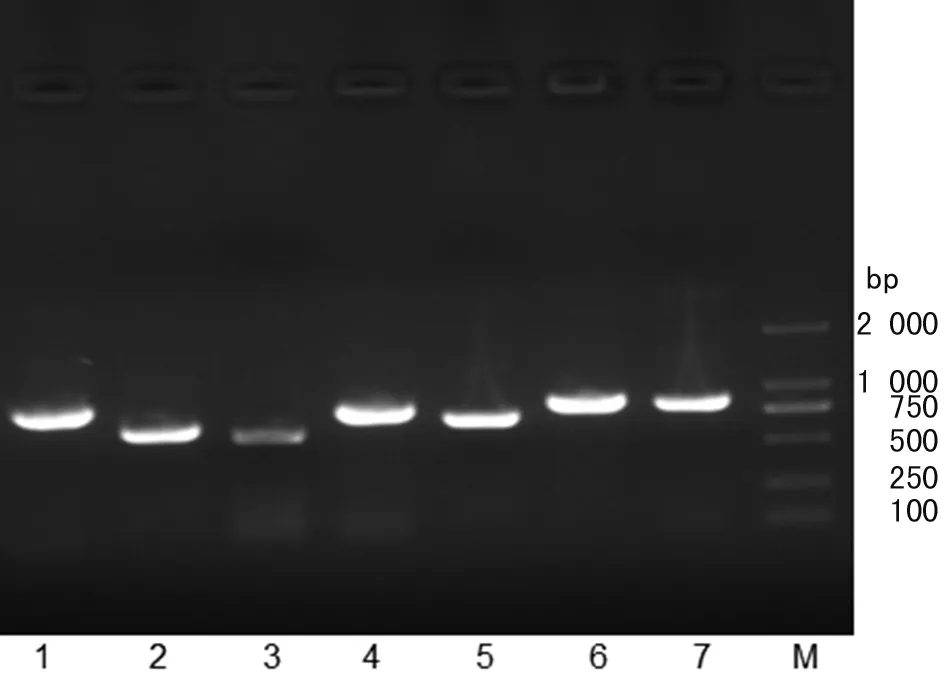
1.thrA; 2.sucA; 3.purE; 4.hemD; 5.hisD; 6.dnaN; 7.aroC; M.DNA Marker Trans 2K图1 管家基因的PCR 扩增结果Fig.1 PCR amplification of housekeeping gene
2.4 管家基因的测序结果和血清型结果 如表5和表6所示,鸡源沙门氏菌分为6个血清型和7个ST型,其中以ST11为主,鸭源沙门氏菌分为2个血清型和3个ST型,其中以ST11为主,猪源沙门氏菌分为3个血清型和3个ST型,其中以ST34为主。总体来看,血清型的分型数目和ST型数目基本相当,这说明MLST与血清学分型的分型能力相近,两者有着紧密的联系。但是,MLST分型的分型更加详细。
从表3-8中我们可以发现,鸡源的沙门氏菌无论是血清型还是ST型,其在总数上都是分布范围最广的。此外,将分离菌的分型结果和其分离地区联系起来,发现同一地区和同一养殖场所采样品的分离菌株的血清型和ST型基本一致。同时,实验还发现了两种新的ST型,分别命名为New1和New2(表7)。
表5 不同动物源的菌株ST型和血清型数量的对比
Tab.5 Comparison between ST type and number of serotype

型别Type鸡源chicken鸭源duck猪源swine血清型serotype623ST型STtype733
表6 不同动物源的菌株ST型和血清型数量的对比
Tab.6 Comparison between ST type and number of serotype
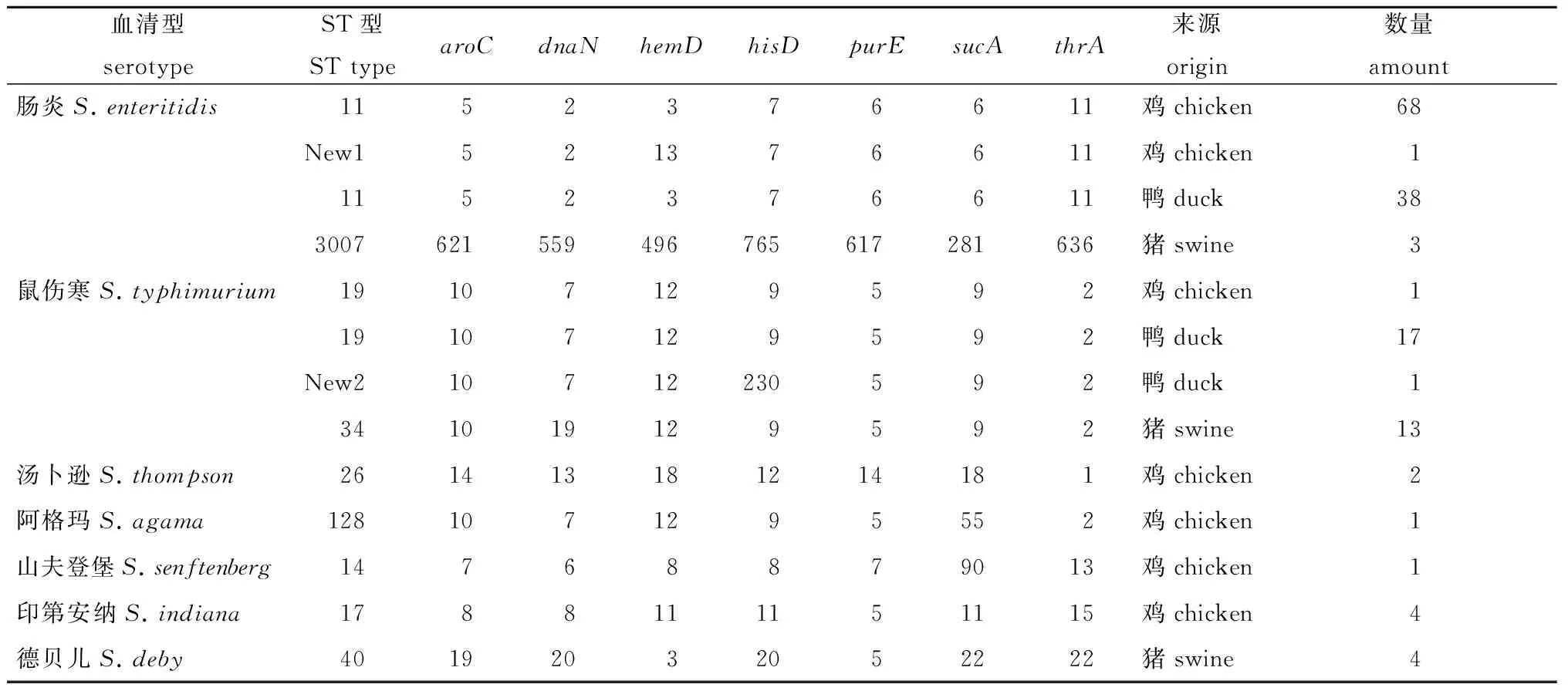
血清型serotypeST型STtypearoCdnaNhemDhisDpurEsucAthrA来源origin数量amount肠炎S.enteritidis1152376611鸡chicken68New1521376611鸡chicken11152376611鸭duck383007621559496765617281636猪swine3鼠伤寒S.typhimurium19107129592鸡chicken119107129592鸭duck17New210712230592鸭duck1341019129592猪swine13汤卜逊S.thompson261413181214181鸡chicken2阿格玛S.agama1281071295552鸡chicken1山夫登堡S.senftenberg14768879013鸡chicken1印第安纳S.indiana1788111151115鸡chicken4德贝儿S.deby40192032052222猪swine4
表7 各地区ST型分布
Tab.7 ST type of different regions
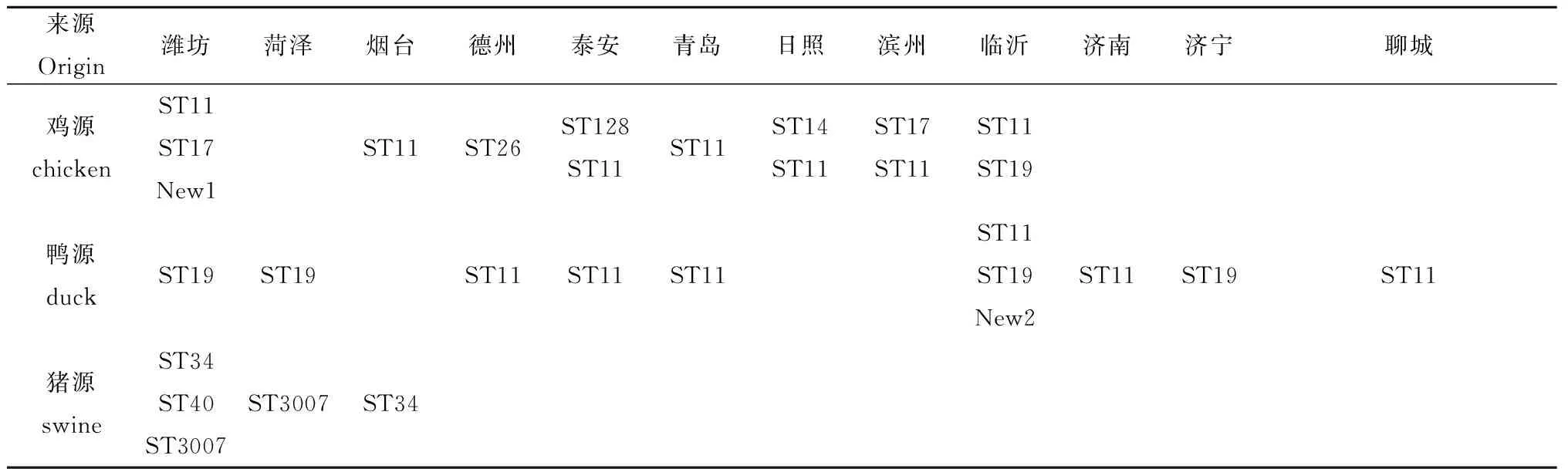
来源Origin潍坊菏泽烟台德州泰安青岛日照滨州临沂济南济宁聊城鸡源chickenST11ST17New1ST11ST26ST128ST11ST11ST14ST11ST17ST11ST11ST19鸭源duckST19ST19ST11ST11ST11ST11ST19New2ST11ST19ST11猪源swineST34ST40ST3007ST3007ST34
3 讨论与分析
3.1 流行病学特点 目前全球报道的沙门氏菌血清型达2 500种以上,我国已报道了 292种不同的血清型,分属于35个O群[10]。而不同区域养殖的模式不同,沙门氏菌的分型特点也存在差异。
本文通过大范围采样检测,表明山东省肠炎沙门氏菌、鼠伤寒沙门氏菌占采样的98%,鸡鸭源肠炎沙门氏菌是主要流行菌株,猪源鼠伤寒沙门氏菌比例最多,而肠炎沙门氏菌的数量则较少,种属特点很明显。鸡源沙门氏菌血清型和ST型远多于鸭,鼠伤寒沙门氏菌、印第安纳沙门氏菌、汤卜逊沙门氏菌、阿格玛沙门氏菌和山谷登堡沙门氏菌均是少量存在,说明鸡源沙门氏菌变异较大。汤卜逊沙门氏菌是在死亡鸡胚中分离出的,而在1994年就有研究人员[11]从死亡的鸡胚中分离出来了汤卜逊沙门氏菌。
王贵升等[12]在山东地区调查小规模养鸡场沙门氏菌的流行分布,发现沙门氏菌70%为副伤寒沙门氏菌,同时也存在鸡白痢沙门氏菌。陈培荣等人[13]在皖北地区调查发现肉种鸡和蛋鸡养殖场分离沙门氏菌的血清型主要是鸡白痢沙门氏菌、肠炎沙门氏菌和鼠伤寒沙门氏菌,其中鸡白痢沙门氏菌占 57.6%,肠炎沙门氏菌占36.0%。这可能与种鸡、商品鸡以及流行阶段的特点有关。
鸭源的沙门氏菌中以肠炎和鼠伤寒沙门氏菌为主要流行菌株,文献中关于山东地区鸭源沙门氏菌的分型报告比较少,钟传德[14]等人对从中国各地区分离的鸭源沙门氏菌进行血清分型,发现肠炎型沙门氏菌61.8%,鼠伤寒沙门氏菌占31.9%;余晓龙等[15]从四川7个地区规模化鸭场分离鉴定到的48株沙门氏菌,其中鼠伤寒沙门氏菌占37.5%和肠炎型沙门氏菌占14.6%。皆与本文结果相似。
猪源沙门氏菌则以鼠伤寒沙门氏菌为主要流行菌株,德贝儿和肠炎沙门氏菌也存在少量的分布。这与刘鲜鲜等[16]在山东地区调查猪屠宰场屠宰环节沙门氏菌污染状况时,发现德尔卑沙门氏菌、鼠伤寒沙门氏菌和汤卜逊沙门氏菌为主要的血清型也有相同之处,此外M Denis[17]等人从种猪和育肥猪场分离的沙门氏菌的血清型主要是德尔卑沙门氏菌和鼠伤寒沙门氏菌,这说明不仅山东地区或国内,乃至国外的猪源沙门氏菌血清型分布都有相似性。
上述比较说明沙门氏菌存在种属和区域的流行大趋势,也因为当地的气候和养殖环境存在明显的差异,是指导区域性防治的重要依据。
3.2 MLST与血清分型的比较 多位点序列分型(MLST)作为一种常用的适合流行病学调查研究的方式,除了具备传统方法的优点,还可以更确切地进行种群的结构以及生物进化方面的研究[18]。如表5,鸡源沙门氏菌分为6个血清型和7个ST型,鸭源沙门氏菌分为2个血清型和3个ST型,猪源沙门氏菌分为3个血清型和3个ST型,存在密切的对应关系:比如鸡鸭ST11对应的血清型是肠炎沙门氏菌,ST19为鼠伤寒沙门氏菌,ST14为山夫登堡沙门氏菌;猪ST40则为德贝儿沙门氏菌等。依据细菌的基因组特征和表型特征的不同来进行的分型有着高度的相关性。
如果仅从两种分型的数量上看,两种分型方式并没有特别大的差距,但是通过Simpson指数(D值)可以评价两种分型方法的能力,其中血清学分型方法D值为0.93,MLST分型方法D值为0.94,两者结合在一起D值则为0.95。
究其原因,沙门氏菌在适应动物内环境和外环境的过程中,ST管家基因的点突变,导致ST型变化,如鼠伤寒沙门氏菌的ST19在鸡鸭一致,而在hisD序号变化,则出现New2,但是鸭宿主特异性依然没有改变;而在管家基因dnaN这一个管家基因的序号不同,导致猪和禽宿主差异性ST34;而当各个管家基因都不同时,才形成新的血清型,如鸡的小比例血清型。肠炎沙门氏菌也存在同样的规律,但是不同宿主ST的管家基因变化已经面目全非了。另外,从地域看(表6),不同地域之间的ST型的分布也有很大差距,说明在传播过程中,沙门氏菌也在进行着进化变异。
两种分型的结合能够提供沙门氏菌流行感染的溯源和进化依据,揭示变异本质。
[1] Poppff MY. Supplement 1998(no.42) to the Kauffmann white scheme[J]. Res Microbiol, 2000, 151(1): 63-65. DOI: 10.1016/S0923-2508(00)00126-1
[2] Sun HL, Jiang Y, Li SC, et al. Analysis of serotype, biofilm-forming ability and antimicrobial resistance ofSalmonellastrains isolated form animals[J]. Acta Vet Et Zootech Sin, 2012, 43(10): 1630-1638. (in Chinese)
孙化露,姜逸,李树纯.动物源性沙门氏菌的血清型、生物被膜形成能力和耐药性分析[J].畜牧兽医学报,2012,43(10):1630-1638.
[3] Chen J, Jiang WC, Tan T, et al.Salmonellapathogenicity island and type III secretion system[J]. Chin J Zoonoses, 2015, 31(4): 371-376. (in Chinese)
陈俊,蒋文灿,谭天,等. 沙门氏菌毒力岛及III型分泌系统研究进展[J]. 中国人兽共患病学报,2015, 31(4): 371-376.
[4] Liu X, Xu YM, Wang B, et al. Application of multi-locus sequence typing (MLST) in the identification ofEscherichiaalbertii[J]. Chin J Zoonoses, 2015, 31(11): 1033-1036. (in Chinese)
刘祥,许彦梅,王斌,等. 多位点序列分型(MLST)在艾伯特埃希菌鉴定中的应用[J]. 中国人兽共患病学报, 2015, 31(11): 1033-1036.
[5] Lian K, Ye SC, Yin YL, et al. Applications of molecular typing methods to the epidemiology ofListeriamonocytogenes[J]. Chin J Zoonoses, 2015, 31(8): 757-762. (in Chinese)
连凯,叶舒畅,殷月兰,等. 分子分型方法在单核细胞增生性李斯特菌流行病学研究中的应用[J]. 中国人兽共患病学报,2015, 31(8): 757-762.
[6] Liu ZG, Wang M, Liu RH, et al. Multilocus sequence typing ofBrucellaclinical isolated strains in Inner Mongolia[J]. Chin J Zoonoses, 2016, 32(11): 991-996. (in Chinese)
刘志国,王妙,刘日宏,等. 内蒙古布鲁氏菌临床分离株多位点序列分型研究[J]. 中国人兽共患病学报,2016, 32(11): 991-996.
[7] Maiden MC, Bygraves JA, Feil E, et al. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms[J]. Proc Natl Acad Sci U S A, 2008, 102(95): 3140-3145. DOI: 10.1073/pnas.95.6.3140
[8] Hanninen ML, Hakkinen M, Rauteli H. Stability of related human and chickenCampylobacterjejunigenotypes after passage through chick intestine studied by pulsed-field gel electrophoresis[J]. Appl Environ Microbiol, 1999, 65(5): 2272-2275.
[9] Steinbrueckner B, Ruberg F, Kist M. Bacterial genetic fingerprint: a reliable factor in the study of the epidemiology of humanCampylobacterenteritis[J]. J Clinic Microbiol, 2001, 39(11): 4155-4159. DOI: 10.1128/JCM.39.11.4155-4159.2001
[10] Peng HB, Wu DF, Kong FD, et al. Distribution of salmonella contamination situation of our country[J], Chin J Front Health Quarant, 2006, 29(2): 125-128. (in Chinese)
彭海滨,吴德峰,孔繁德,等.我国沙门氏菌污染分布概况[J].中国国境卫生检疫杂志,2006, 29(2): 125-128.
[11] Bao LD, Tian YM, Tu BD, et al. PathogenicE.coliandSalmonellaseparation and identification from chicken embryos[J]. Inner Mong Vet, 1994, (4): 44-45. (in Chinese)
宝力德,田有明,图布丹扎布,等.鸡胚致病性大肠埃希氏菌及沙门氏菌的分离及鉴定[J].内蒙古兽医,1994,0(4):44-45.
[12] Wang GS, Tian FL, Chen J, et al. Epidemiological investigation of salmonellosis in some small-scale chicken farm of Shandong area[J]. Poul Sci, 2007, 0(5): 11-13. (in Chinese)
王贵升,田夫林,陈静,等. 山东省小规模养鸡场沙门氏菌流行病学调查分析[J]. 家禽科学,2007,0(5):11-13.
[13] Chen PR, Liu JN, He B. Isolation and identification of avianSalmonellafrom north of Anhui and antibiotic resistance research[J]. Agr Sci Hubei, 2013, 52(17): 4161-4163. (in Chinese)
陈培荣,刘锦妮,何斌. 皖北地区禽源性沙门氏菌分离鉴定及耐药性研究[J]. 湖北农业科学,2013,52(17):4161-4163.
[14] Zhong CD. Isolation, Identification, drug resistance profile investigation and primary study on plasmid characteristic of Pathogenic salmonella from poult duck in some cities and provinces of China[D]. Ya’an: Sichuan Agricultural University, 2006. (in Chinese)
钟传德. 我国部分省市雏鸭致病性沙门氏菌分离鉴定、耐药谱调查及携带质粒特性的初步研究[D].雅安:四川农业大学,2006.
[15] Yu XL. Studies on the isolation, analysis of serotypes, drug susceptibility and drug resistant genes ofSalmonellafrom ducks Sichuan[D]. Ya’an: Sichuan Agrigultural University, 2012. (in Chinese)
余晓龙.四川地区鸭源沙门氏菌分离与血清型分析及其药敏性与耐药基因相关性研究[D].雅安:四川农业大学, 2012.
[16] Liu XX, Wang JW, Wang J, et al. Analysis onSalmonellacontamination status during pig slaughtering in Shandong province[J]. Chin Animal Health Inspect, 2015, 32(5): 82-86. (in Chinese)
刘鲜鲜,王君玮,王娟,等. 山东部分生猪屠宰场屠宰环节沙门氏菌污染状况分析[J]. 中国动物检疫,2015,32(5):82-86.
[17] Denis M, Houard E, Fablet A, et al. Distribution of serotypes and genotypes ofSalmonellaentericaspecies in French pig production[J]. Vet Rec, 2013, 173(15): 370-370.
[18] Liao YL. Pathogenic microorganisms MLST research progress[J]. Progr Microbiol Immun, 2007, 35(4): 65-68. (in Chinese)
廖亚玲.病原微生物基因多位点序列分型的研究进展[J].微生物学免疫学进展,2007,35(4):65-68.
Distributionandtypingofanimal-derivedSalmonellawithMLSTandserotype
ZHAO Cui1, ZHANG Qing2, GUO Shu-yuan3, LI Lu-lu2, LIU Yu-jie4, WANG Yong-ming5, CHANG Wei-shan3, LIU Yu-qing2
(1.Tai’anCityAnimalHealthSupervisionCenterinShandongProvince,Tai’an271000,China;2.AnimalHusbandryandVeterinaryInstitute,ShandongAcademyofAgriculturalScience,Jinan250100,China; 3.AnimalScienceAcademyofShandongAgriculturalUniversity,Tai’an271018,China;4.AnimalEpidemicPreventionandControlCenterinRizhaoCity,Rizhao276800,China; 5.ShandongHuaHongBiologicalProductsCo.,Ltd.,Binzhou256600,China)
To compare the discriminatory ability of multilocus sequence typing and serotype of animal-derivedSalmonellaand find its distribution in Shandong Province, 78 chicken-origin, 56 duck-origin and 20 swine-originSalmonellawere separated from some regions of Shandong Province. Seven conserve sequences ofSalmonellawere PCR-amplified for MLST and slide agglutination test for serotyping. Results showed that by serotyping, 6 serotypes were identified from chicken-originSalmonella, including 88.5%S.enteritidis, 5.1%S.indiana, 2.6%S.thompson, 1.3%S.typhimurium, 1.3%S.senftenberg, 1.3%S.agama. Two serotypes were identified from duck-originSalmonella, including 67.9%S.enteritidis, 32.1%S.typhimurium. Three serotypes were identified from swine-originSalmonella, including 65%S.typhimurium, 20%S.derby, and 15%S.enteritidis. By MLST typing, seven ST types were identified from chicken-originSalmonella: ST11, ST19, ST26, ST128, ST14, ST17 and New1. Three ST types were identified form duck-originSalmonella: ST11, ST19 and New2.
Three ST types were identified from swine-originSalmonella: ST34, ST40 and ST3007. Overall, the types identified with two methods were closed, so MLST and serotype have similar discriminatory ability.
Salmonella; serotype; MLST; typing Supported by the Shandong Provincial Major Projects of Independent Innovation, Shandong Dhicken Public Health Model Establishment and Demonstration, Shandong Academy of Agricultural Science and Technology Innovation Project (No. CXGC2016A10)
Liu Yu-qing, Email: liuiuqing@163.com
1.山东省泰安市动物卫生监督所,泰安 271000; 2.山东省农业科学院畜牧兽医研究所,泰安 250100; 3.山东农业大学动物科学院,济南 271018; 4.山东省日照市动物疫病预防控制中心,日照 276800; 5.山东华宏生物工程有限公司,济南 256600
R183.1
:A
:1002-2694(2017)09-0793-07
2017-03-13编辑:刘岱伟
山东省农业科学院科技创新工程(CXGC2016A10),山东省农业重大应用技术创新项目(SDAIT-11-09)
通迅作者:刘玉庆,Email: liuiuqing@163.com
猜你喜欢
首都食品与医药(2022年21期)2022-12-06
动物医学进展(2022年9期)2022-09-06
中国食品学报(2022年6期)2022-07-19
中国饲料(2021年17期)2021-11-02
医药前沿(2021年25期)2021-10-16
食品安全导刊(2021年20期)2021-08-30
今日农业(2021年1期)2021-03-19
养猪(2020年6期)2021-01-27
保健文汇(2020年11期)2020-12-03
河南畜牧兽医(2020年11期)2020-01-11
