
城市绿地中臭椿更新苗空间点格局及种间关联分析
2017-09-26 07:05万媛媛李洪远贺梦璇
水土保持通报 2017年4期
万媛媛, 李洪远, 贺梦璇
(南开大学 环境科学与工程学院, 天津 300350)
城市绿地中臭椿更新苗空间点格局及种间关联分析
万媛媛, 李洪远, 贺梦璇
(南开大学环境科学与工程学院,天津300350)
[目的] 探究城市绿地中臭椿更新苗的数量特征、存活情况及空间分布格局,为城市绿地生境下乔木的天然更新和人工干扰或促进更新提供理论依据。 [方法] 运用种群静态生命表,点格局分析法中的双关联函数G(r)、H(r)函数分析臭椿更新苗的生存状况,空间分布格局及各龄级间的空间关联性。 [结果] 更新苗种群结构存在波动性,存活曲线趋于DeeveyⅡ型,低龄级和中龄级个体数量丰富,个体死亡率和消失率在前期和后期较稳定,中期先减后增,超过第5龄级(8~10 cm)后,个体死亡率和消失率出现高峰。 [结论] 一侧和两侧铺装样地在空间上都呈现出不同程度的聚集分布,聚集强度随尺度距离的增大呈减小趋势,一侧铺装样地在尺度接近3.7 m处有转为随机分布的趋势。龄级间,小尺度内多表现为无关联,随着尺度增加呈正显著相关;两侧铺装有正关联的龄级组合比一侧铺装样地多;较大尺度上,用径级代表龄级的更新苗间相关关系越接近,正关联越强。
城市绿地; 臭椿; 更新苗; 空间点格局; 空间关联性
文献参数: 万媛媛, 李洪远, 贺梦璇.城市绿地中臭椿更新苗空间点格局及种间关联分析[J].水土保持通报,2017,37(4):79-84.DOI:10.13961/j.cnki.stbctb.2017.04.014; Wan Yuanyuan, Li Hongyuan, He Mengxuan. Spatial point pattern and specific association of regenerated saplings ofAilanthusaltissimain urban greenland[J]. Bulletin of Soil and Water Conservation, 2017,37(4):79-84.DOI:10.13961/j.cnki.stbctb.2017.04.014
空间点格局分析起源于植物生态学研究,由于克服了单一尺度下种群分布格局分析的不足,还可以对种群的水平结构给以定量的描述而被广泛应用。目前关于空间分布格局在大尺度森林群落种群中的研究较多,主要集中在森林群落中优势物种的多尺度空间分布格局、优势物种的不同龄级以及不同物种间的多尺度空间关联研究等方面[1-6],而对小尺度城市绿地,城市森林更新种群幼苗的生长状态和空间分布状态的研究很少。种群种间关系,分布特征与空间格局的研究紧密关联[7]。种内联结用来表示同一物种不同龄级在空间分布上的相互关联性,是对一定时期内植物种群组成个体之间相互关系的静态描述,这种关系不仅包括空间分布关系,也包含着个体间的功能关系,揭示了种群的现状,并能反映种群内个体间的相互作用[8-9]。在自然界中,每一种生物都具有一定的空间分布型,甚至同一种生物在不同生长发育阶段以及不同的生境下其分布型也存在较大差异[10-11]。幼苗期是对环境条件最为敏感的时期,是决定植物种群能否天然更新的重要阶段[12]。因此,研究城市绿地中臭椿幼苗的生存状况和更新格局,有助于补充和丰富人们对城市生境下乔木天然更新状况的了解。另外,幼树幼苗的分布格局能帮助理解成年植株的生长发展和更新潜力,为城市绿地生境下乔木的天然更新和人工干扰或促进更新提供理论基础和科学依据。
1 研究区概况及研究方法
1.1 样地设置
在天津市南开区水上公园,动物园和南开大学校园3个公园绿地分别设置样地,以铺装条件作为生境因子,将待调查样地分为无铺装(Ⅰ类),一侧铺装(Ⅱ类)和两侧铺装(Ⅲ类)3类,一侧铺装,铺装类型为水泥道路,两侧铺装一侧为楼房建筑,一侧为水泥道路。各样地选取的母树株数分别为2棵(Ⅰ类)、2棵(Ⅱ类)和3棵(Ⅲ类),母树平均胸径分别为32.65,37.05和35.8 cm,胸径相差不大,年龄级相当,基本为同期栽植,具有代表性;3类样地草本层地被伴生种主要为早熟禾,泥胡菜,灰菜,地被植物组成相似,种类较单一。样地内,以选定母树为起点,采用相邻格子法,将每个样地划分为多个1 m×1 m的相邻网格,统计每一个格子内的更新苗的数量,以道路铺装和更新苗出现的最远距离为标准划定方形的样地范围,样地面积为340 m2(Ⅰ类)、150 m2(Ⅱ类)、280 m2(Ⅲ类),同时记录更新苗地径、高度、距母树的距离以及坐标方位,样地内植被特征,地形和地貌形态。
1.2 数据处理
1.2.1 种群径级划分 根据实际调查数据,剔除无法记录位置信息或地径级别极小的更新苗后,将样地内更新苗地径和高度进行分类,Ⅰ—Ⅲ类样地内更新苗数分别为207,92,191棵,更新苗高度以2 cm为极差,划分为12个级别,进行种群静态生命表编制(见表1);样地内更新苗地径普遍较小,因此,将地径级别划分如下:地径级1(0,1 mm],地径级2(1,2 mm]直至地径级8(7,8 mm],调查中没有发现更新苗地径在(8,9 mm]间的记录,因此处理时,将地径级9代表的范围定为(9,10 mm],一侧铺装样地中没有发现地径级别为1的幼苗,两侧铺装样地地径级最高到第8径级。
1.2.2 种群静态生命表编制 生命表是研究种群数量动态的一个重要手段,生命表和存活曲线不仅可以反映种群生存现状,还可以展现植物种群与环境适应的结果,对阐明群落未来发展趋势也具有重要意义[13]。采用“空间代替时间”法编制[14],用高度级结构代替年龄结构,由于特定时间生命表反映了多个世代重叠的年龄动态历程中的一个特定时间,而不是对这一种群全部生活史的追踪,并且在调查中存在系统误差,在生命表中会出现死亡率为负的情况。对于这种情况,Wentworth[15]认为,生命表分析中产生一些负值,这与数学假设技术不符,但仍能提供有用的生态学记录,表明种群并非静止不动,而是在迅速发展或衰落中,为此,采用匀滑技术处理(ax*)(表1)。为了进一步对臭椿种群更新苗生存状况进行分析,特引入生存分析中的4个函数项目于生命表栏目中,计算公式为
Sx=P1·P2·P3,…,Px
(1)
Fx=1-Sx
(2)

(3)
λx=fx/Sx=2qx/〔hx(1+px)〕
(4)
式中:Sx——生存函数;Px——存活频率;Fx——积累死亡率函数;fx——死亡密度函数;hx——区间长度;qx——死亡频率;λx——危险率函数[16]。
1.2.3 种群分布点格局 根据臭椿更新苗的空间坐标数据,绘制二维空间分布的点图,以此为基础使用双关联函数G(r)公式(5)进行臭椿更新苗多尺度的种群格局分析。G(r)函数由Ripley的K函数推演而来[17-18],利用点间的距离,计算任一点为圆心,半径为r的指定宽度圆环区域内的点的数量来进行空间点格局分析,与Ripley的K函数相比,G函数能较敏感地判别某一尺度上点的实际分布偏离期望值的程度,是评估聚集程度的重要分析方法(公式5)。用多元点格局的分析方法探讨不同的发育阶段更新苗的格局动态及制约机制,用H(r)代替K(r)[19],在随机分布下,K(t)/π可以使方差保持稳定,同时H(r)与r有线性关系,其表达式为:

(5)
K12(r)反映龄级1和龄级2在空间尺度上的关联性,计算以龄级1的每个个体为圆心,半径为r的圆内龄级2的数量。通过Monte Carlo方法模拟了99%置信区间,用种群实际分布数据(点图)计算得到的不同尺度下的G(r),H(r)值,单变量分析中,G(r)在包迹线之间,为随机分布,高于上包迹线为聚集性分布,低于下包迹线为均匀分布;在双变量分析中,H(r)值在包迹线以内,表示2个龄级无显著关联;大于上包迹线时,为显著性正相关,小于下包迹线时,为显著性负相关。点格局分析通过R软件包spatstat实现。
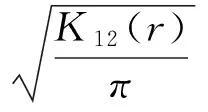
(6)
2 结果分析
2.1 臭椿更新苗种群生命表与存活曲线
根据静态生命表的编制方法和无铺装样地(自然状态下)更新苗高度级的调查数据,编制臭椿更新苗种群的静态生命表并估算生存函数,在第1,3,5龄级的数据发生波动,小于第2,4,6龄级,发生异常。据特定生命表假设,年龄组合是稳定的,各年龄的比例不变。因此,将数据分为2个区段:1~6龄级和8~12龄级。分别计算2个区段存活数的累积:
(7)
(8)
平均数分别为ax1=T1/n1=186/6=31,ax2=T2/n2=9/5=1.8≈2,且认为这2个平均数是区段的组中值。另外,根据2个区段的最多存活数和最少存活数的差数(分别为40,7)及区段的间隔数量(6,5),可以确定2个区段相邻年龄组的存活数之间的差约为7,1左右。故经匀滑修正后,得到ax*(表1)。幼苗种群结构存在波动性Ex反映了x龄级内个体的生命期望寿命,总体期望值较低,最高值仅为0.212,并随着龄级的增加而减小。存活曲线是一条借助于存活个体数量来描述特定年龄存活率。臭椿更新苗随着龄级增加,成活率呈下降趋势,在第6龄级的下降趋势稍缓(图1),存活曲线明显趋于DeeveyⅡ型(直线);死亡率(qx)和消失率(Kx)曲线变化趋势基本一致(图1),在第5龄级有明显的下降趋势,第6龄级种群大量减少,种内和种间竞争最剧烈,但特征值总体波动不大,波动范围在0.1~0.45左右;种群生存率单调下降,累计死亡率单调上升,二者互补(图1);种群的死亡密度曲线和危险率曲线在第2龄级后开始出现较明显反差,特征值均较大,死亡密度曲线在0.5及以下基本持平,第6龄级死亡密度最大,危险率曲线总体呈递增趋势,在第5~7龄级呈现急剧增长。

表1 臭椿种群静态生命表及生存函数估算
注:ax表示存活数;ax*表示匀滑修正值;lx表示存活量;Lx表示区间寿命;dx表示死亡量;qx表示死亡率;Tx表示总寿命;Ex表示期望寿命;Kx表示消失率;Sx表示生存率函数;Fx表示累计死亡率函数;fx表示死亡密度函数;λx表示危险率函数。

图1 臭椿更新苗种群存活曲线与生存分析函数
2.2 臭椿更新苗种群空间分布
臭椿更新苗种群的个体分布点图如图2所示,图中星号为母树位置。点格局分析的空间尺度多为样地最小边长的1/2,本文结合道路铺装距离和更新苗出现的最远距离,选择经R软件计算推荐的空间参考尺度,即:一侧铺装和两侧铺装样地分别为0~3.75 m和0~3.30 m。无铺装样地更新苗定位数据不完整,因此根据记录的一侧铺装和两侧铺装样地更新苗位置以及地径数据,进行点格局分析。通常在小尺度上形成的空间分布格局主要是受种群自身生物学特性的影响,而在较大尺度上的空间分布格局则更多的是受到生境异质性的作用。从种群个体分布的点图上看,两类样地在空间上都呈现出不同程度的聚集分布。两侧铺装样地在0~3.3 m范围内均为聚集分布,曲线位于包迹线上方,距离包迹线距离较远,随着距离的增加,聚集程度下降;一侧铺装样地整体呈聚集分布,只在极小尺度(0~0.04 m)和在接近3.7 m处曲线位于包迹线内侧偏上位置,表现出随机分布,中间距离均为聚集分布,有时极小尺度的随机分布距离太小可以忽略。一侧和两侧铺装样地呈现出了较一致的空间分布趋势,即在研究尺度范围内以聚集性分布为主,但随着研究尺度的增大呈现随机分布。

注:❋为母树位置。
2.3 更新苗不同发育阶段的空间关联
臭椿更新苗在两侧铺装样地0~3.3 m尺度上的种间关系分析,不同年龄级(地径级)的组合共有28种,各大小龄级间的空间关联(包括正负关联和无关联3种关联情况)结果基本表现为5大类,如表2所示,A型表现为无显著相关;B型表现为正显著相关且随距离增加正相关性逐渐增大;C型为正显著相关,随距离增加正显著相关性有减小的趋势;D型为正显著相关且1~3.3m尺度范围内, 相关性不稳定或正显著相关间隔、间断分布;E型表现为小尺度范围内无显著相关性,大尺度呈正显著相关。各龄级总体上在较小的尺度上,曲线多被上下包迹线包围,空间关联微弱;在0~3.3 m模拟范围内随着尺度的增大,H(r)值接近或大于包迹线的机会或程度更大。0.33 m空间范围内有10个龄级组合(表2)表现为无关联,龄级组合数占总组合数比例最大为35.71%(A型);0~0.5 m外的实际曲线在包迹线上,随距离增加分别增加/减小的龄级组合,占总组合数比例分别为17.86%(B型)和21.43%(C型);龄级组合1~2,1~5,2~6间的空间相关性规律性不明显,正相关性在距离上是间隔分布的,在0~2.5 m出现多个细小区段的显著正相关,划分在D型;龄级组合5~7,1~3,1~4,5~6在0~2 m范围内(较小的距离区段内)无显著相关性,超过一定的距离范围后(中等距离范围),随着空间距离的增大呈现正相关,即聚集分布,幼苗的空间分布有一定代表性,因此划分在E型。总体表现为具有相关关系的龄级组合所占比例较大,为64.29%,占组合数的1/2以上,且都为正相关。一侧铺装样地,龄级间多数无显著相关关系(82.14%),空间关联曲线被包在包迹线范围之间,如2~5,2~6组合比较明显。
一侧铺装样地无E型相关性组合,B型龄级组合2~3在小尺度(0~1 m)范围内无显著空间相关,随着距离的增加,正相关越来越显著,尺度内相关性最大可达到2;C型龄级组合2~4和组合3~4的正显著相关性随距离增加有下降的趋势;龄级组合4~5和组合5~7总体呈显著正相关,属于D型。但进行多次Monte Carlo模拟,正相关区域多间隔出现,规律性不明显。龄级间,两侧铺装比一侧铺装样地正关联种对较多,一侧铺装样地空间内有显著正相关的在两侧铺装样地也成正显著相关;正相关径级多集中在第2~5级的小径级之间。

表2 更新苗龄级组合及其空间关联
3 讨 论
3.1 臭椿幼苗种群结构与动态
城市绿地中更新苗种群的存活和死亡情况对研究更新苗种群结构、更新动态、更替规律、种群和环境之间的关系以及其未来发展与演变趋势有重要的意义。种群生命表分析表明,低龄级臭椿更新苗数量储备丰富,主要集中在龄级1~2,但幼树数量不足,且生命期望值随着年龄的增加而减小,除第1龄级外,生命期望总体维持在0.1左右,更新苗成活率较低,样地的幼苗死亡率非常高,有的甚至是刚出苗不久就全部死亡。存活曲线与生存分析函数图表明,更新苗种群实际数量在第3龄级,第9~11龄级出现明显的瓶颈现象,说明经过环境和自身筛选,只有少数幼苗能顺利进入第3和第12两个较高龄级;但死亡率在第5龄级表现最低,生存率明显提高,且死亡高峰出现在相邻的第6龄级。Augspurger等发现,多数风播树种的种子在光照和荫蔽条件下都能萌发,但多数种的幼苗在光照下比在荫蔽下生长好,一年之后幼苗的生长高度也高于荫蔽下的幼苗[20]。本研究中,同样是中间龄级(第5龄级)的幼苗存活率高,说明随着树龄的增大,空间和资源环境较适宜其生长,超过第5龄级,土壤、光照和水分条件不能满足所有幼苗生长需要,种内竞争增加,空间生态位受到限制,自疏和他疏作用加强。因此,在更新苗种群发育到第5龄级时,有选择地采取干扰(进行人工稀疏),有利于更新苗种群的生长和存活。
3.2 种群分布格局
种群空间格局分析是研究种群特征、种间关系以及种群与环境之间相互关系的重要手段[21]。张金屯曾指出,点格局的取样面积以正方形或矩形样地为宜,边长应该在50 m以上[1]。对于植物更新苗,由于个体较小,往往零星分布或呈斑块状,如果取样面积过大,很可能在运用点格局法时会由于样地面积太大而导致结果更多地反映斑块的空间特征,从而掩盖了植物种群不同个体的空间格局特征[2]。通常认为在较小的尺度上,不同的分布格局由生物学特性决定,如种内、种间竞争,种子扩散限制,幼苗补充过程等;在更大的尺度上,则是由环境的异质性造成的,如水分、光照、土壤养分和温度等因素。臭椿本身喜光,不耐阴,不同生长阶段对光照强度的要求不同。本研究中,两种样地尺度连续变化条件下,臭椿幼苗极小范围内表现为随机分布,随着距离的增加呈聚集分布,且种群格局和格局强度随尺度的增加而减小,研究结果与孙志虎等[22]研究长白山红松林天然林树木在幼林或苗期阶段呈明显的聚集分布的研究结果相似,集群分布与种子传播方式、发生以及周围的环境密切相关,推测光辐射分布的不均匀是不同空间位置上更新苗生长存在差异和更新格局为聚集分布的一个重要原因;不同生境条件的样地间,0~6 m空间范围内,两侧铺装样地比一侧铺装样地空间聚集强度更大,表明两侧铺装样地对空间的侵占程度较高,推测可能是由于人为扰动带来的种子迁移。城市绿地铺装形式及比率复杂多样,难以找到同样条件的重复样地,本研究以现有绿地为研究对象,并非按照试验要求设计绿地形式和铺装比例以增加重复和对照。
3.3 种间关联性
种群在幼年阶段较高的集群强度有利于群体效应的发挥,促进种群的存活和发展[23],本研究中,两侧铺装样地,在2~3.3 m尺度范围内,第1龄级和第2,3,4,5龄级间有显著的正相关关系,随着龄级的增加空间相关性随之增加,龄级越大的更新苗的空间分布关系与最小径级更新苗的空间关联性越大,越相互吸引,结果与刘炜洋等无干扰条件下水曲柳幼苗高大植株与幼小植株空间关联性下降的研究结果相反;但一侧铺装样地的龄级2,两侧铺装样地的龄级3,4和龄级5与稍大龄级个体间空间关系,表现为相邻龄级空间正相关较大,推测更新苗个体受相邻龄级更新苗个体影响较大,可能是由于还没有密度制约的条件下,相邻龄级幼苗有相同的环境和资源的需要,这样吸引的聚集的分布还可以提高该物种共同抵抗病菌等不适合生存条件侵害,提高物种整体的存活率[24]。
4 结 论
(1) 臭椿更新苗种群结构存在波动性,存活曲线趋于DeeveyⅡ型,低龄级和中龄级个体数量丰富,个体死亡率和消失率在前期和后期较稳定,中期先减后增,中间龄级(第5龄级8~10 cm)的幼苗存活率较高,第6龄级个体死亡率和消失率出现高峰。
(2) 一侧和两侧铺装样地在空间上臭椿幼苗极小范围内表现为随机分布,随着距离的增加呈聚集分布,聚集强度有随尺度距离的增大呈减小趋势,一侧铺装样地在尺度接近3.7 m处有扩散趋势,转为随机分布;
(3) 龄级组合,两侧铺装的正关联龄级组合比一侧铺装样地多;龄级间,多在小尺度内无关联,随着尺度增加呈正显著相关;第1龄级与龄级越大的更新苗的空间分布的关联性越大,越相互吸引;较大尺度上,2个龄级更新苗间相关关系越接近,正关联越强。
[1] 张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[2] Wiegand T, Moloney K A. Rings, circles and null-models for point pattern analysis in ecology[J]. Oikos,2004,104(2):209-229.
[3] 刘保双,付登高,吴晓妮,等.滇中次生常绿阔叶林优势树种的空间格局[J].生态学杂志,2013,32(3):551-557.
[4] 任珩,赵成章.高寒退化草地狼毒与赖草种群空间格局及竞争关系[J].生态学报,2013,33(2):435-442.
[5] Olagoke A O, Bosire J O, Berger U. Regeneration of Rhizophora mucronata(Lamk.)in degraded mangrove forest: Lessons from point pattern analyses of local tree interactions[J]. Acta Oecologia, 2013,50(7):1-9.
[6] 李伟,王瑞雪,张光富,等.南方红豆杉迁地保护种群的点格局分析[J].生态学杂志,2014,33(1):16-22.
[7] 黄明钗,史艳财,韦霄,等.珍稀濒危植物金花茶的点格局分析[J].生态学杂志,2013,32(5):1127-1134.
[8] 高福元,赵成章,卓马兰草.高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性[J].生态学报,2014,34(16):4698-4706.
[9] 王磊,孙启武,郝朝运,等.皖南山区南方红豆杉种群不同龄级立木的点格局分析[J].应用生态学报,2010,21(2):272-278.
[10] Thai V T. The Rain Forest Ecosystem of Vietnam[M]. Hanoi: Science Technology Press, 1998:321-345.
[11] Pham Van huong, 陈昌雄,洪伟,等.不同林分下胖大海幼树幼苗更新分布格局[J].森林与环境学报,2015,35(4):377-384.
[12] 王妍,卢琦,吴波,等.呼伦贝尔沙地樟子松种群更新潜力[J].浙江农林大学学报,2011,28(2):248-253.
[13] 冯金朝,袁飞,徐刚.贵州雷公山自然保护区秃杉天然种群生命表[J].生态学杂志,2009,28(7):1234-1238.
[14] 吴承祯,洪伟,吴继林,等.珍稀濒危植物长苞铁杉的分布格局[J].植物资源与环境学报,2000,9(1):31-34.
[15] Wentworth T R. Laboratory and field exerises in ecology[J]. Ecology, 1982,63(6):1987.
[16] 卢杰,潘刚,罗大庆,等.濒危植物急尖长苞冷杉种群生命表分析[J].水土保持通报,2010,17(5):212-221.
[17] Ripley B D. The second-order analysis of stationary point processes[J]. Journal of Applied Probability, 2004,13(2):255-266.
[18] Illian J, Penttinen A, Stoyan H, Stoyan D. Statistical Analysis and Modelling of Spatial Point Pat-terns[M]. US: John Wiley & Sons Ltd.,2008.
[19] 刘炜洋.水曲柳天然更新及其影响因子研究[D].黑龙江 哈尔滨:东北林业大学,2008.
[20] 张巧明,辽东栎林天然更新特征的研究[D].陕西 杨凌:西北农林科技大学,2007.
[21] Greig S P. Quantitative Plant Ecology[M].3rd ed. London: Blackwell Scientific Publications,1983:19-128.
[22] 孙志虎,张彦东.长白落叶松人工林天然更新幼苗分布格局及其研究方法的比较[J].生物数学学报,2009,24(3):556-566.
[23] 蔡飞,宋永昌.武夷山木荷种群结构和动态的研究[J].植物生态学报,1997,21(2):138-148.
[24] 彭闪江,黄忠良,彭少麟,等.植物天然更新过程中种子和幼苗死亡的影响因素[J].广西植物,2004,24(2):113-121.
Spatial Point Pattern and Specific Association of Regenerated Saplings ofAilanthusAltissimain Urban Greenland
WAN Yuanyuan, LI Hongyuan, HE Mengxuan
(College of Environmental Science and Engineering, Nankai University, Tianjin 300071, China)
[Objective] The quantitative characteristics, the survival condition and the spatial distribution pattern ofAilanthusaltissimaregenerated saplings were investigated and analyzed to provide theoretical bases for natural regeneration of arbor, human-induced disturbance regeneration, and other regeneration promotion in urban green space habitat. [Methods] Time-specific life table, paired correlation functionG(r) andH(r) were utilized to analyze the seedlings’ living conditions, the distribution pattern and spatial association among seedlings with different age classes. [Results] There was a fluctuation in the structure ofAilanthusaltissimaregenerated saplings’ population. The individuals at young and middle age-class were abundant, and the type of survival curve of the population was regarded as Deevey Ⅱ. Lifespan’s curve of mortality rate and disappearance rate all had a stable stage in the early and late period, while in the middle period, the two items experienced decrease and increase successively. Individual mortality and disappearance rates reached the top peaks beyond the 5th age-class(8~10 cm). [Conclusion] Regenerated saplings were significantly aggregated in different degree and the aggregation intensity weakened with the increase of scale in both two pavement’s types. The type of one side’s pavement seemed randomly distributed when the scale is close to 3.7 m. There is almost no correlation relationship at small scale, however, when scale increased, positive correlation relationships between seedlings of different age classes was detected. The closer the age class of the regenerated saplings was, the stronger the positive correlation was.
urbangreenland;Ailanthusaltissima;regeneratingsaplings;spatialpointpattern;spatialassociation
A
: 1000-288X(2017)04-0079-05
: Q145
2016-11-02
:2017-01-26
国家自然科学基金项目“不同生境下城市绿化树种VOCs排放影响机制及估算模型研究”(31370700)
万媛媛(1991—),女(汉族),天津市人,硕士研究生,主要从事恢复生态学研究。E-mail:wanyuanyuan@mail.nankai.edu.cn。
李洪远(1963—),男(汉族),天津市人,教授,博士生导师,主要从事植被恢复与生态重建、土壤种子等方面的研究。E-mail:econklee@126.com。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
基层中医药(2021年4期)2021-07-22
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
天津农林科技(2018年1期)2018-03-07
运动(2017年1期)2017-09-03
湖北农业科学(2017年11期)2017-07-13
广西农学报(2015年4期)2015-09-11
东方女性(2014年10期)2014-11-05
东北林业大学学报(2014年8期)2014-08-02
