
水曲柳天然林种群的点格局分析1)
2014-08-02 03:54:12许彦明张彦东孙海龙
东北林业大学学报 2014年8期
许彦明 张彦东 孙海龙
(东北林业大学,哈尔滨,150040)
刘炜洋
(黑龙江省双鸭山市林业局)
水曲柳天然林种群的点格局分析1)
许彦明 张彦东 孙海龙
(东北林业大学,哈尔滨,150040)
刘炜洋
(黑龙江省双鸭山市林业局)
在黑龙江省五常市宝龙店林场,利用点格局分析法分析了水曲柳(Fraxinusmandshurica)种群不同龄级的空间分布格局和空间关联性。结果表明:龄级Ⅰ、Ⅱ、Ⅳ在0~50 m尺度上呈聚集分布,龄级Ⅲ在0~21 m尺度上呈聚集分布,21~50 m尺度上呈随机分布。水曲柳种群各龄级的数量随龄级增加呈倒“J”型分布,龄级Ⅰ个体数量最高,占总数量的89.5%,由龄级Ⅰ到龄级Ⅱ个体数量骤减,而龄级Ⅱ到龄级Ⅳ个体数量逐渐减少。龄级Ⅰ个体与其他龄级空间关联性较小,龄级Ⅱ与龄级Ⅲ、龄级Ⅲ与龄级Ⅳ的空间关联性总体较小,龄级Ⅱ与龄级Ⅳ在0~17 m尺度上呈负关联,17~50 m尺度上关联性较小。这说明龄级Ⅰ个体在空间上受其他龄级的影响较小,而龄级Ⅱ个体在空间上受到了龄级Ⅳ个体较大的影响。
水曲柳;点格局分析;空间分布格局;空间关联性
By point pattern analysis, we studied the spatial distribution pattern and the spatial association of different age class Manchurian ash (Fraxinusmandshurica) population in Baolongdian Forest Farm, Wuchang, Heilongjiang Province. The age class I, II and IV had a clumped distribution at a scale 0-50 m. Age class III had a clumped distribution at a scale 0-21 m and a random distribution at a scale 21-50 m. The amount of each age class of ash population was a reverse “J” shape distribution with the increasing of age class. The amount of age class I individual is the highest, accounting for 89.5%. The amount decreased strongly from age class I to age class II, but the amount decreased slowly from age class II to IV. The spatial association between age class I and other three age classes at 0-50 m was feeble, as well as between age class II and III, and between age class III and IV in the mass. Between age class II and IV, the spatial association was significantly negative at 0-17 m, but feeble at the scale 17-50 m. An individual in age class I was less affected by other age class in space, but individual in age class II was greatly affected by age class IV in space.
种群空间格局是指种群个体在水平空间的分布状况,通常种群空间分布格局与物种的生物学特性、种内和种间竞争及环境条件等多种因素密切相关[1]。种群分布格局与尺度密切相关,长期以来人们提出了多种分析种群格局的方法,但到目前为止,点格局分析能很好的分析多种尺度下种群的格局[2-3]。通过点格局分析,可以从多尺度上了解植物种群的空间分布格局和种间关联性,以及种群与环境之间的关系[3-4]。
水曲柳(Fraxinusmandshurica)是东北林区的重要珍贵用材树种,因其材质优良长期以来被过度采伐利用,目前该树种资源已濒于枯竭[5]。为更好地培育水曲柳后备资源,人们开展了水曲柳种群更新与动态问题的探讨。稍早的研究表明,水曲柳是东北东部山地具有较大更新优势的树种,在多数群落中水曲柳天然更新均较好[6],尤其在针叶树人工林下水曲柳天然更新具有明显优势[7-10]。不过近年的研究表明,尽管水曲柳在多种群落中具有更新优势,但其在自身林下更新数量相对较少[9-10]。韩有志等[11]研究发现,水曲柳母树附近不利于幼苗的生存,似乎水曲柳母树对自身幼苗更新产生了一定的“排斥”作用。然而目前关于不同龄级水曲柳个体间关系的研究尚不多见,为此,本文利用点格局分析法分析水曲柳天然种群不同龄级的分布格局及龄级间的关联性,探讨不同龄级间的相互影响,并为水曲柳天然林的经营与管理提供理论依据。
1 研究区概况
研究地点选择在黑龙江省五常市宝龙店林场(N44°50′~44°58′,E127°38′~127°55′),该林场位于长白山北部的张广才岭西坡,地貌类型为低山丘陵。该区属温带大陆性季风气候,冬季干燥寒冷,夏季温暖多雨。年降水量约620 mm,且主要集中于6—8月,年蒸发量1 094 mm。年平均气温约3.4 ℃,≥10 ℃积温为2 000~2 500 ℃,无霜期120 d左右。
该地区地带性顶极群落为阔叶红松林,但经过反复破坏(采伐、火烧、开垦等)后,目前形成以水曲柳、山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、蒙古栎(Quercusmongolica)、黄檗(Phellodendronamurense)和胡桃楸(Juglansmandshurica)等为主要树种的次生林。地带性土壤为暗棕壤,多分布于山地斜坡,非地带性土壤有白浆土、草甸土和沼泽土,主要分布于丘陵漫岗和河岸两侧的低地。
2 研究方法
2.1 样地设置与调查
研究林分是以水曲柳为主的天然次生林,林分树种株数组成为:水曲柳72.5%,白榆(Ulmuspumila)12.6%,白桦6.7%,胡桃楸5.6%,山杨2.1%,以及少量的黄檗和五角槭(Acermono)。于2007年7月在林分内设置100 m×100 m的样地,用相临网格法将样地划分为100个10 m×10 m的小样方。在每个小样方内,逐株测定水曲柳个体的胸径(D),并记录水曲柳个体的位置以坐标值表示。由于实际测定树木年龄比较困难,所以本文以树木胸径大小作为表征树木年龄大小的指标。根据调查数据并参考其他研究[3],将水曲柳种群按胸径大小划分为4个龄级,龄级Ⅰ:D<2.5 cm,并包括树高不足1.3 m的幼苗幼树;龄级Ⅱ:2.5 cm≤D<10.0 cm;龄级Ⅲ:10.0 cm≤D<20.0 cm;龄级Ⅳ:D≥20.0 cm。
2.2 点格局分析方法
2.2.1 种群各龄级空间分布格局分析
密度(λ)和协方差(k)是二维数集的一次和二次特征结构。对于点格局,λ是单位面积内的期望点数,k是点间距离分布的测定指标,k随着尺度的变化而变化。Diggle证明该二次特征结构可以简化为一个函数方程K(r),公式[3]为:
K(r)=λ-1。
(1)
式中:K(r)为表征二次特征结构的随机分布函数,r为研究尺度,可以是大于0的任何值;λ为单位面积内的期望点数,可以用n/A来估计,A为样地面积,n为总点数(植物个体数)。
在实践中,K(r)用下式估计[3]:
(2)
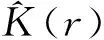
(3)
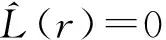
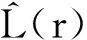
2.2.2 种群龄级间的空间关联性分析

(4)
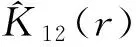
(5)

仍用Monte-Carlo检验拟合包迹线,以检验两个龄级间是否呈显著关联性。
2.3 数据分析
利用ADE—4软件对水曲柳不同龄级种群进行点格局分析。本文采用的最大距离尺度为样地边长的一半(50 m),步长值为1 m,Monte-Carlo模拟检验100次,得到由上下2条包迹线围成的99%的置信区间。采用Sigmaplot 10.0软件绘制格局图形。
3 结果与分析
3.1 水曲柳种群结构
在所调查的林分内,水曲柳种群各龄级间数量波动较大,随龄级增加呈倒“J”型分布。龄级Ⅰ的数量为4 481株/hm2,而龄级Ⅱ、龄级Ⅲ和龄级Ⅳ数量分别仅为:269、137和122株/hm2(图1),龄级Ⅰ的数量明显高于其他龄级,占总数量的89.5%。由龄级Ⅰ到龄级Ⅱ数量骤减,减幅达94.0%,但从龄级Ⅱ到龄级Ⅲ、龄级Ⅲ到龄级Ⅳ的数量逐渐减少,分别减少49.1%和10.9%。龄级Ⅰ、龄级Ⅲ在样地内分布较均匀,龄级Ⅱ、龄级Ⅳ主要分布在样地的两侧。

图1 水曲柳各龄级个体在样地中的分布点图
3.2 水曲柳种群各龄级空间分布格局
3.3 水曲柳种群各龄级间空间关联性
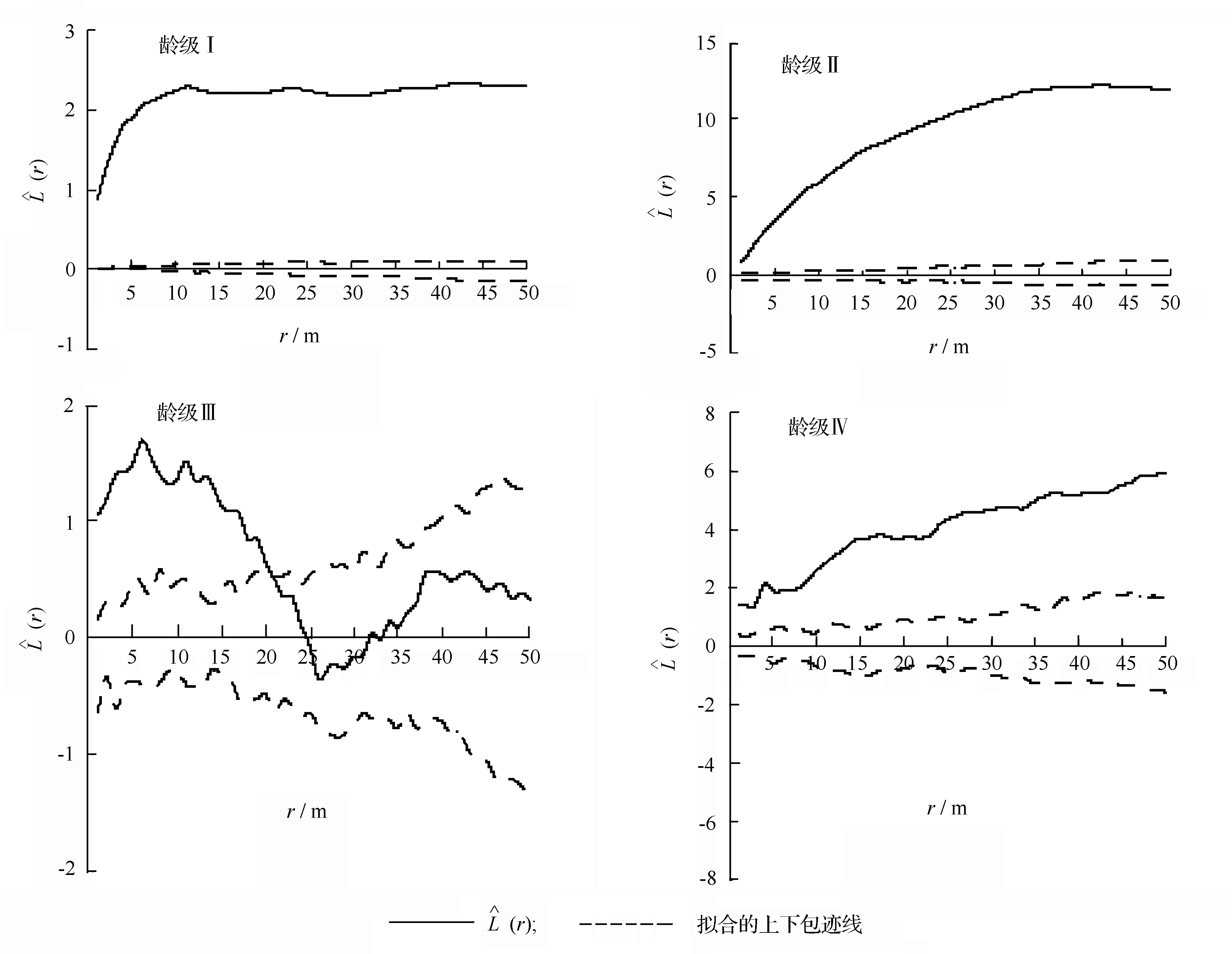
图2 水曲柳各龄级的空间分布格局
4 结论与讨论
在我们所调查的林分中,水曲柳种群龄级Ⅰ、Ⅱ、Ⅳ在0~50 m尺度内呈现聚集分布,龄级Ⅲ在0~21 m尺度上呈现聚集分布,21~50 m尺度上呈现随机分布。多数研究表明,植物以母株为中心的繁殖方式使低龄个体呈聚集分布,随着年龄的增加,树木对光因子和营养条件的要求加大而发生自然稀疏等作用,种群会趋于均匀分布,随着年龄的继续增加,种群将趋于随机分布[12-13]。本研究龄级Ⅰ、龄级Ⅱ呈聚集分布,龄级Ⅲ在一定尺度上由聚集分布逐渐趋于随机分布这与多数研究结果相似[12,14-15],而龄级Ⅳ在所研究尺度内呈聚集分布与多数研究结果不同,这可能与该林分前期受到择伐干扰有关。因择伐时砍伐了部分大径级的个体,使得连续的冠层在空间上出现空隙[16],从而使保留下来的龄级Ⅳ个体表现为聚集分布的格局。
水曲柳种群龄级Ⅰ个体数量为4481株/hm2,但到龄级Ⅱ阶段个体数量骤减到269株/hm2。这说明从龄级Ⅰ到龄级Ⅱ阶段水曲柳幼苗幼树大量死亡,这与以前的一些研究结果是一致的[17-18]。类似的现象在其他树种的研究中也曾被观察到[19-21]。进一步的空间关联性分析表明,水曲柳龄级Ⅱ个体与龄级Ⅳ个体二者在0~17 m尺度上达到空间负关联,说明水曲柳龄级Ⅱ个体在生长过程中受到了附近龄级Ⅳ个体的竞争排斥,这可能是导致水曲柳龄级Ⅱ个体存活数量较少的主要原因。不过在本研究中水曲柳龄级Ⅰ个体与其他龄级间在所研究尺度上空间关联性较小,这似乎说明在本研究林分中龄级Ⅰ个体并未受到其他龄级个体较大的影响。然而韩有志等[11]研究发现,在水曲柳母树集群分布区,更新幼苗较少,母树附近不利于自身幼苗的生存。还有一些研究也证明,水曲柳幼苗在自身林下更新表现不良[9-10],本研究的水曲柳龄级Ⅰ个体数量为4 481株/hm2,更新亦属较少,这与以前的研究结果相似[10]。柏广新等[10]的研究认为,水曲柳林下自身更新不良可能与种内的他感作用有关。本研究出现水曲柳龄级Ⅰ个体与其他龄级间空间关联性较小,可能与所研究林分母树(龄级Ⅲ和龄级Ⅳ)分布密度较高有关(259株/hm2)。在水曲柳母树密度相对较高的林分中,分布在母树间不同位置的龄级Ⅰ个体可能普遍受到来自母树的竞争或他感作用影响,而且距母树不同位置的龄级Ⅰ个体受到的影响程度较接近,故导致龄级Ⅰ个体在林地上分布相对较均匀,表现出与其他龄级的空间关联性较小。而韩有志等研究的林分母树密度较低(12 hm2样地内5株母树),在母树间不同位置的幼苗可能受到来自母树的竞争或他感作用压力差异较大,远离母树位置的压力小,而母树附近受到的压力大,故导致更新幼苗在空间分布上表现出母树附近较少的现象。

图3 水曲柳各龄级间的空间关联性
[1] Dale M T. Spatial pattern analysis in plant ecology[M]. Cambridge: Cambridge University Press,1999:207-276.
[2] Dale M T, Powell R D. A new method for characterizing point patterns in plant ecology[J]. Journal of Vegetation Science,2001,12(5):597-608.
[3] 张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35-40.
[4] 刘振国,李镇清.不同放牧强度下冷蒿种群小尺度空间格局[J].生态学报,2004,24(2):227-234.
[5] 孔冬梅,沈海龙,冯丹丹,等.水曲柳体细胞胚与合子胚发生的细胞学研究[J].林业科学,2006,42(12):130-133, 封3.
[6] 王义弘,柴一新,慕长龙.水曲柳的生态学研究[J].东北林业大学学报,1994,22(1):1-6.
[7] 沈海龙,丁宝永,夏辛畅,等.人工针叶群落中水曲柳种群发生发展特点的研究[J].东北林业大学学报,1991,19(S1):182-190.
[8] 王承义,刘关彬.人工林下天然更新种群特征[J].林业科技,2000,25(6):13-15.
[9] 刘炜洋,陈国富,张彦冬.不同林分内水曲柳天然更新及影响因子研究[J].华东森林经理,2010,24(4):19-23.
[10] 柏广新,张彦东.水曲柳天然更新及其影响因子[J].东北林业大学学报,2013,41(1):7-9, 13.
[11] 韩有志,王政权.天然次生林中水曲柳种子的扩散格局[J].植物生态学报,2002,26(1):51-57.
[12] Laaka-Lindberg S, Hedderson T A, Longton R E. Rarity and reproductive characters in the British hepatic flora[J]. Lindbergia,2000,25(2/3):78-84.
[13] 郭华,王孝安,肖娅萍.秦岭太白红杉种群空间分布格局动态及分形特征研究[J].应用生态学报,2005,16(2):227-232.
[14] 吴大荣,朱政德.福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究[J].林业科学,2003,39(1):23-30.
[15] 缪宁,史作民,冯秋红,等.川西亚高山岷江冷杉种群的空间格局分析[J].林业科学,2008,44(12):1-6.
[16] Rigg L S. Disturbance processes and spatial patterns of two emergent conifers in New Caledonia[J]. Austral Ecology,2005,30(4):363-373.
[17] 王广发,庄发明,赵俊昌.长白山林区水曲柳人工更新方式的研究[J].吉林林业科技,2006,35(3):13-18.
[18] 张健,李步杭,白雪娇,等.长白山阔叶红松林乔木树种幼苗组成及其年际动态[J].生物多样性,2009,17(4):385-396.
[19] Packer A, Clay K. Soil pathogens and spatial patterns of seedling mortality in a temperate tree[J]. Nature,2000,404:278-281.
[20] Nathan R, Safriel U N, Noy-Meir I, et al. Spatiotemporal variation in seed dispersal and recruitment near and far from Pinus halepensis trees[J]. Ecology,2000,81(8):2156-2169.
[21] Gilbert G S, Harms K E, Hamill D N, et al. Effects of seedling size, El Nio drought, seedling density, and distance to nearest conspecific adult on 6-year survival of Ocotea whitei seedlings in Panamá[J]. Oecologia,2001,127(4):509-516.
Point Pattern Analysis of Manchurian Ash (Fraxinusmandshurica) Natural Population/
Xu Yanming, Zhang Yandong, Sun Hailong
(Northeast Forestry University, Harbin 150040, P. R. China);
Liu Weiyang
(Forestry Bureau of Shuangyashan City, Heilongjiang Province)//Journal of Northeast Forestry University.-2014,42(8).-40~45
Manchurian Ash (Fraxinusmandshurica); Point pattern analysis; Spatial distribution pattern; Spatial association
1) 国家科技支撑计划课题(2012BAD21B0202-02);中央高校基本科研业务费专项(DL10BA02)资助。
许彦明,男,1987年8月生,东北林业大学林学院,硕士研究生。
张彦东,东北林业大学林学院,教授。E-mail:zhyd63@163.com。
2013年12月19日。
S718.5
责任编辑:潘 华。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
贵州林业科技(2021年4期)2021-12-23 12:14:16
山东林业科技(2021年4期)2021-08-31 05:55:04
林业科技情报(2021年1期)2021-04-14 09:10:02
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
农民致富之友(2019年11期)2019-05-23 01:14:52
森林工程(2018年4期)2018-08-04 03:23:10
福建林业科技(2018年1期)2018-04-09 00:56:41
农民致富之友(2017年23期)2018-01-02 02:02:06
