
豆腐在不同储藏温度下腐败菌菌相比较
2017-09-07 00:13:48吴丽樱成玉梁郭亚辉孙秀兰钱和
食品研究与开发 2017年18期
吴丽樱,成玉梁,郭亚辉,孙秀兰,钱和
(江南大学食品学院,江苏无锡214122)
豆腐在不同储藏温度下腐败菌菌相比较
吴丽樱,成玉梁,郭亚辉,孙秀兰,钱和*
(江南大学食品学院,江苏无锡214122)
以盒装内酯豆腐和简易包装老豆腐为研究对象,包括这两种豆腐的新鲜样品和在不同温度储藏后(4、25、37℃)的腐败样品,提取样品细菌基因组后对16S rRNA V3区基因片段扩增,采用MiSeq高通量测序技术,分析腐败菌菌相组成及细菌多样性情况。结果表明:盒装内酯豆腐的菌相复杂程度小于简易包装老豆腐,盒装内酯豆腐中的细菌主要由芽孢杆菌属(Bacillus)组成,比例均在90%以上。简易包装老豆腐在4℃腐败后相比其他两个温度下的样品,细菌多样性更丰富。明串珠菌属(Leuconostoc)是25℃和37℃下腐败老豆腐的主要优势菌,分别占到了71%和80%。高通量测序相比于传统的可培养技术,反映的微生物信息更加接近于真实的样品生态,此研究可为小型豆腐生产企业的卫生控制提供参考。
盒装内酯豆腐;简易包装老豆腐;高通量测序;腐败菌菌相
豆腐在我国是深受百姓喜爱的传统美食,价格低廉,美味可口,又能提供丰富的蛋白质及多种功能成分,在日常膳食中有着不可取代的地位[1]。目前中国的豆制品生产基本仍以地方性小企业、小作坊为主,存在生产规模小,卫生状况差,自动化程度低等问题[2]。豆腐制品含水量高,营养丰富,为微生物提供了理想的繁殖条件,极易腐败变质,阻碍了豆制品行业向规模化、产业化发展。
目前对于豆腐中腐败菌的研究大都基于传统的可培养技术,其结果不能很好地还原样品中菌相的真实状态。而新兴的二代测序技术能够大规模且全面地解析微生物多样性,在发酵[3]、土壤[4-5]、医学[6]等各个领域都发挥了重要作用,彻底革新了微生物生态学研究,但该技术在腐败菌研究中的运用还较少。
盒装内酯豆腐和简易包装老豆腐是生活中最常食用的两种豆腐,由于生产工艺和包装条件的不同(生产工艺如图1),两种豆腐的腐败菌相也有所差别。为了更好地了解其菌相差异,本文采用MiSeq测序分析了同一工厂生产的盒装内酯豆腐和简易包装老豆腐,分别在新鲜状态,以及4、25、37℃下储藏直到腐败后微生物菌群多样性,加深对导致豆腐腐败的微生物的认识,同时对豆腐企业的工艺改进和质量控制具有指导意义。
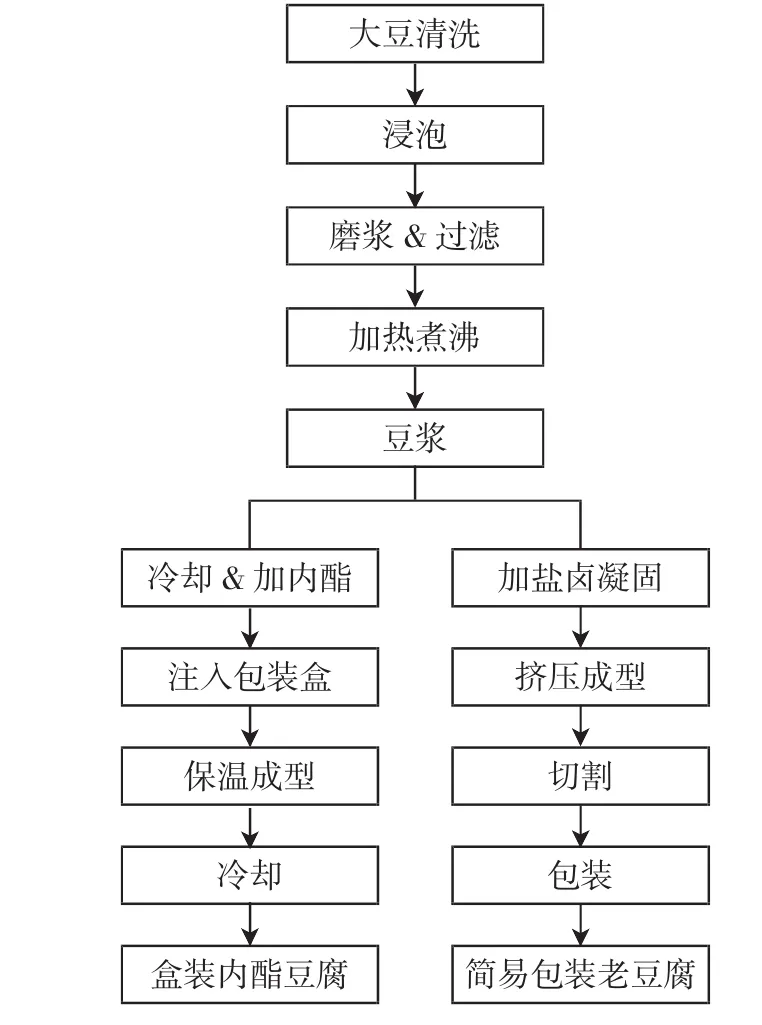
图1 两种豆腐生产工艺流程图Fig.1 The process flow diagram of two kinds of tofu
1 材料与方法
1.1 材料与仪器
简易包装老豆腐和盒装内酯豆腐:无锡香道嘉食品厂;DNA提取试剂盒E.Z.N.A.Soil DNA Kit:OMEGA公司;Qubit2.0 DNA检测试剂盒:Life公司;Taq DNA Polymerase:Thermo 公司;Agencourt AMPure XP:Beckman公司。
Pico-21 台式离心机:Thermo Fisher;TND03-H-H漩涡混合器:海门市其林贝尔仪器制造有限公司;TND03-H-H混匀型干式恒温器:深圳拓能达科技有限公司;DYY-6C型电泳仪电源、DYCZ-21电泳槽:北京市六一仪器厂;Biodoc-IT凝胶成像系统:美国UVP公司;T100TMThermal Cyeler PCR仪:BIO-RAD公司。
1.2 方法
1.2.1 样品的准备
将豆腐厂刚下线的简易包装老豆腐和盒装内酯豆腐装于有冰袋的保温箱中,2 h之内运回实验室。将部分新鲜样品存放于-80℃冰箱以便后续提取DNA。其余样品,分别置于4、25、37℃下储藏直至腐败(50%感官评价小组成员判定其达到感官不可接受状态)。分别用NF、LF表示新鲜的盒装内酯豆腐和简易包装老豆腐样品;N4、N25、N37分别表示盒装内酯豆腐在4、25、37 ℃下贮藏腐败后的样品;L4、L25、L37 分别表示简易包装老豆腐在4、25、37℃下贮藏腐败后的样品。8个样品的储藏时间和样品名称见表1。
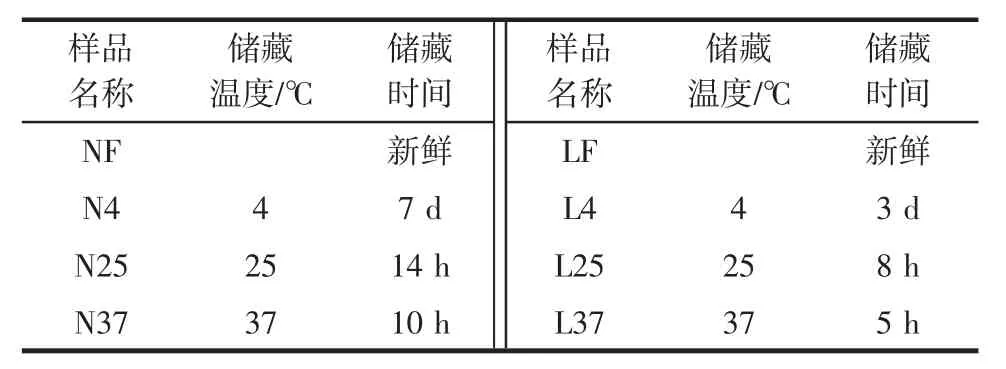
表1 样品名称和储藏时间Table 1 The sample name and storage time
1.2.2 豆腐中细菌DNA的提取
将每一样品取3个平行,称取20 g豆腐样品混匀,取2 mL液体加入无菌离心管中,10 000 r/min温离心3 min,取上清具体提取步骤参照OMEGA试剂盒E.Z.N.ATMMag-Bind Soil DNA Kit的试剂盒使用说明书。将同一样品的3个平行提取的DNA样品进行混合。
1.2.3 16S rRNA V3区基因片段的扩增及Illumina双末端测序
对于提取的基因组进行V3区的扩增,采用引物[7]341F:5’-CCCTACACGACGCTCTTCCGATCTGCC TACGGGNGGCWGCAG-3’,534R:5’-ACTGGAGTTCCTTGGCACCCGAGAATTCCAGATTACCGCGGCTGCTGG-3’。PCR反应体系见表2。

表2 V3区基因片段扩增的PCR反应体系Table 2 PCR reaction system for the amplification of V3 region gene fragment
PCR反应程序如下:94℃预变性3 min;94℃变性30 s,45℃退火20 s,65℃延伸30 s(5个循环);94℃变性20 s,55℃退火 20 s,72℃延伸30 s(20个循环);72℃最终延伸10 min;10℃保温。使用2%浓度的琼脂糖凝胶对PCR产物进行电泳检测。DNA纯化回收,利用Qubit2.0 DNA检测试剂盒对回收的DNA精确定量,按照1∶1的等量混合后测序。每个样品DNA量取10 ng,上机测序浓度为20 pmol。
1.2.4 生物信息学分析
1)测序完成后,为了保证信息分析质量,必须对其进行质控和过滤,将低质量的片段舍弃。使用Usearch去除非扩增区域序列,校正错误序列,并调用uchime鉴定嵌合体,将去除嵌合体的序列与数据库代表性序列进行blastn比对,在97%的相似水平下进行归类操作得到OTU数(分类操作单元)。2)对8个样品进行Alpha多样性分析,使用Mothur(http://www.mothur.org)计算香农指数(Shannon),菌种丰富度指数(Chao1),辛普森指数(Simpson),覆盖率(Coverage)和ACE等常用的α生物多样性指数,衡量样本中物种的多样性[8]。3)菌群分类学分析:将全部OTU序列在Silva数据库中比对,采用贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并在属水平(genus)统计每个样品的群落组成,不满足条件的序列则被归为unclassified,将OTU进行属的鉴定并作图。4)Beta多样性分析,利用Unifrac计算样本间的距离,分析比较8个样品之间的差别度量。
2 结果与分析
2.1 样品基因组的提取与扩增产物检测
对8个样品用试剂盒提取宏基因组后,对16SrRNA的V3区基因片段进行PCR扩增。琼脂糖凝胶电泳检测后结果如图2。
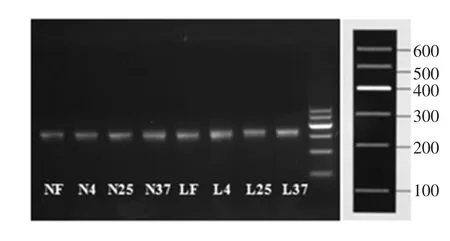
图2 8个样品的V3区扩增产物电泳检测Fig.2 Electrophoresis detection of amplified products in V3 region of 8 samples
由图2可知,目的条带清晰,说明扩增结果良好,产物浓度适宜,可用于后续高通量测序。
2.2 Illumina测序,序列基本信息分析
8个样品的高通量测序基本数据见表3。
如表3所示,8个样品处理后剩余序列在56 000到72 000之间,OTU数在260到590之间。其中,盒装内酯豆腐的4个样品的平均OTU数量约为341,简易包装老豆腐的OTU数目约为461,总体来看,老豆腐的腐败菌种类要多于盒装内酯豆腐。新鲜的内酯豆腐OTU数最少,而其他3个样品的OTU数相近;而老豆腐的样品中,随着储藏温度的升高,样品的OTU数逐渐减少。
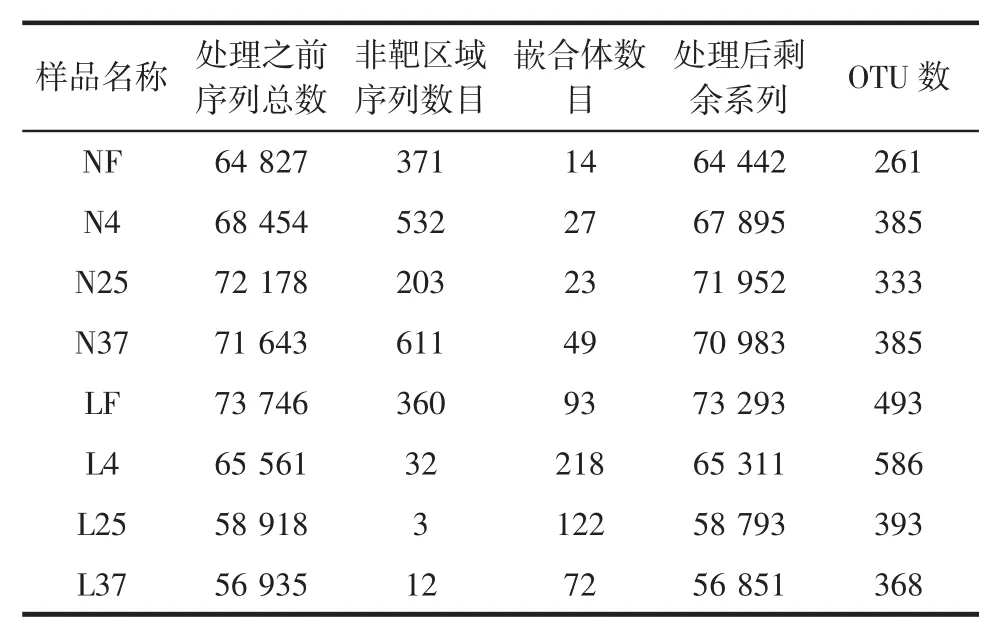
表3 8个样品的高通量测序基本数据Table 3 High-throughput sequencing basic data of 8 samples
2.3 Alpha生物多样性分析
ACE和Chao1在生态学中用来估物种总数,数值越大,代表群落丰富度越高。Shannon和Simpson是反映的是样品中微生物的多样性。Shannon值越大,代表该样品中微生物群落多样性越高,Simpson指数值越大,说明群落多样性越低[9]。
细菌多样性指数见表4。
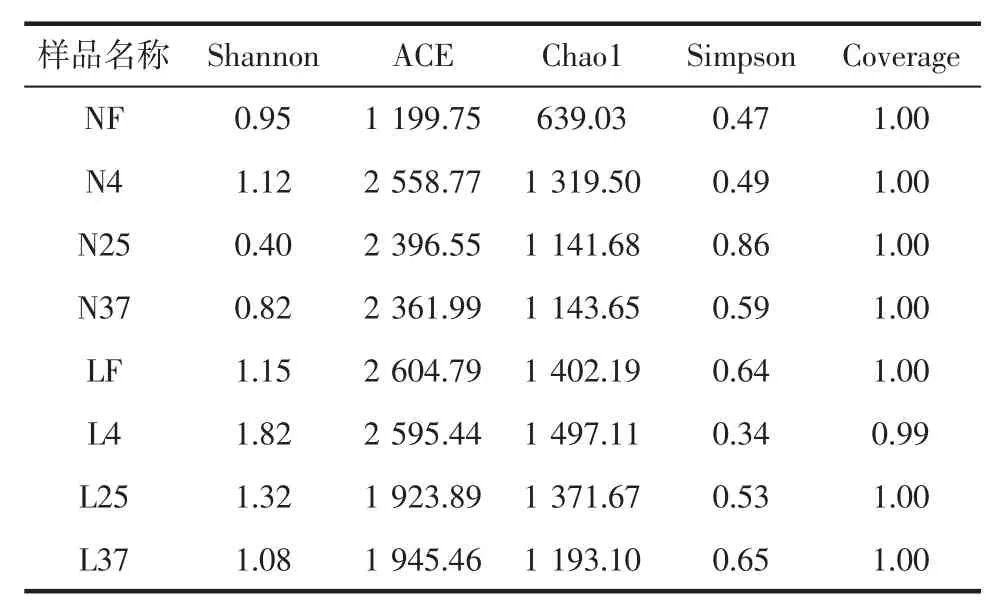
表4 细菌多样性指数Table 4 Alpha-diversity data
由表4可知,老豆腐组的群落多样性明显高于内酯豆腐组,N4和L4样品的群落多样性分别高于同组其他样品,说明在4℃条件下,无论是内酯豆腐还是老豆腐,细菌的种类较25、37℃下更丰富。这可能是由于在低温状态下,大多数的细菌生长缓慢,生理代谢受到了抑制,细菌与细菌之间的竞争减弱,没有因为某种菌的大量生长而造成其他菌的衰弱。这与工商大学薛静[10]的研究一致,她研究了生制章鱼在5、15、25℃的条件下贮藏后腐败菌菌相变化,结果表明在5℃下Shannon最大,生物多样性最高。在老豆腐组中,温度越高,Shannon值越低。当储藏温度升高时,优势菌迅速生长,代谢物质积累,抑制了其他菌的生长,微生物多样性减少。由ACE和Chao1指数可知,N4、N25、N37样品的物种丰富度相似,高于NF样品;而在老豆腐组中,LF和L4样品物种数量相当。Coverage值代表各样本文库的覆盖率,测出的值越接近1,表明各样本覆盖率越高。8个样品的覆盖率均接近1,说明本次测序结果均能很好地反映样本的真实情况。
2.4 豆腐在不同温度储藏腐败后的菌相分析
图3是各个样品在属水平上的细菌丰度图。
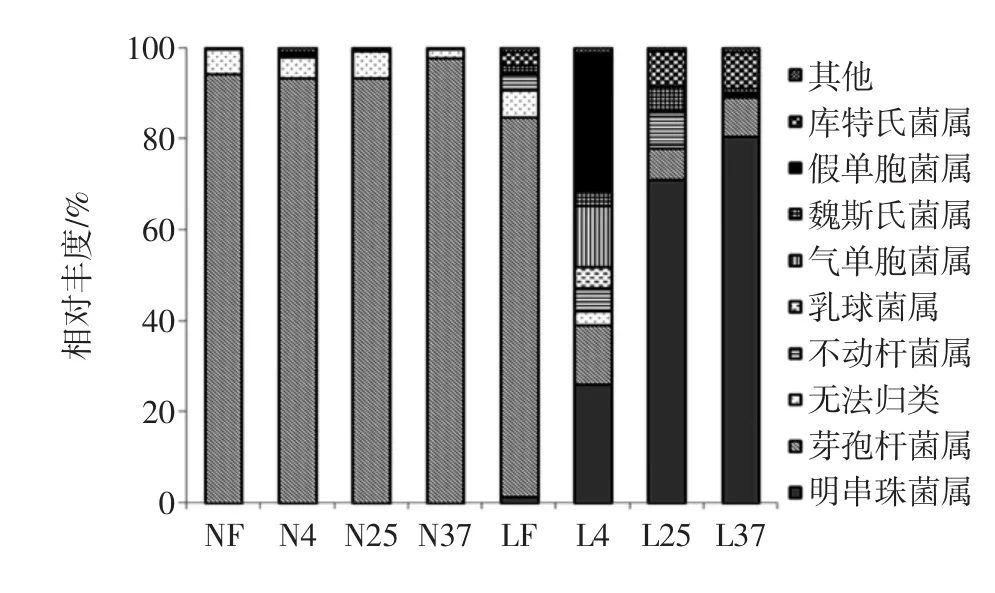
图3 细菌在属水平上的丰度Fig.3 Abundance of bacteria at the genus level
由图3可知,芽孢杆菌属(Bacillus)在内酯豆腐中优势明显,均在90%以上,这与生产工艺有很大关系。内酯豆腐制作过程中有一个保温成型的过程,绝大部分微生物的营养体在此过程中已死亡,只剩下一些耐热的芽孢杆菌的芽孢还存活,后期当条件适宜时这些芽孢又会成为营养体继续繁殖。而老豆腐在凝固后还需要经过挤压成型和切割的过程,除了加热后残留的微生物,还会污染上环境中的各种微生物,所以其初始菌相比内酯豆腐更加复杂。内酯豆腐的初始菌相中,约有94%的芽孢杆菌,还有大概5%的菌归为无法归类,这可能与选取的基因片段有关,无法识别一种或多种细菌,猜测可能是一些其他耐热的细菌,如类芽孢杆菌属(Paenibacillus)。
内酯豆腐中由于大部分不耐热的腐败菌已经在保温成型过程中被杀死,极少部分可能由于中心温度较低,加热不充分而残留,这部分细菌在数量上微乎其微,约占1%以下,再加上生存环境较严苛(密封的豆腐盒中含氧量较少),在后期没有迅速繁殖起来,且运输储藏过程中染菌几率小,因此芽孢杆菌在内酯豆腐的储藏过程中始终占据优势地位。由此可推断,引起盒装内酯豆腐腐败的主要是耐热的芽孢杆菌。这与邓勇[11]、李博[12]等的研究一致。
LF的菌相和内酯豆腐很相似,但是由于后续的工序中污染了环境中或者包装中的其他微生物,芽孢杆菌约占80%。在不同的温度下储藏腐败后,其菌相发生了很大变化。在4℃储藏腐败后下,假单胞菌(Pseudomonas)约占 40%,气单胞菌属(Aeromonas)约占20%,芽孢杆菌属(Bacillus)约12%。假单胞菌属(Pseudomonas)是一种嗜冷菌,尽管在新鲜状态下的豆腐中极其微量存在,但是由于它更适应低温环境,在4℃下有一定程度的增长。假单胞菌产蛋白水解酶能力强,容易在蛋白含量丰富,碳水化合物少的食品中繁殖[13],在腐败的冷藏食品中最常见。气单胞菌(Aeromonas)主要存在于水体,淤泥,土壤和人类粪便中,豆腐中的气单胞菌可能来源于地面溅起的水或者工人的手。低温状态下,芽孢杆菌繁殖能力很弱。明串珠菌属在样品L25和L37中占据了绝对优势,分别为71%,80%。杨明[14]也同样在散装老豆腐中分离鉴定得到了肠膜明串珠菌,反接后证明该菌导致了老豆腐质构变软。明串珠菌属产酸能力强,它的代谢产物使体系的pH值下降,从而抑制了其他菌的生长繁殖。库特氏菌(Kurthia)在L25和L37两个样品中分别占约7%,8%的比例。李除夕[15]对4种不同品牌的豆腐在25℃储藏腐败后同样分离得到两株库特氏菌,并认为库特氏菌属主要通过产生腐胺引起豆腐的腐败气味。L25样品中不动杆菌(Acinetobacter)约占6%,不动杆菌广泛分布于外界环境中尤其是潮湿的环境,主要在水体和土壤中,粘附力极强,易在各类材料上粘附,不动杆菌极易可能来源于豆腐生产的不锈钢设备上,由于接触造成老豆腐的二次污染。
2.5 Beta多样性分析
Beta的数值越接近于0,则表明这样品间在物种多样性方面存在的差异越小。样品间的距离矩阵见表5。
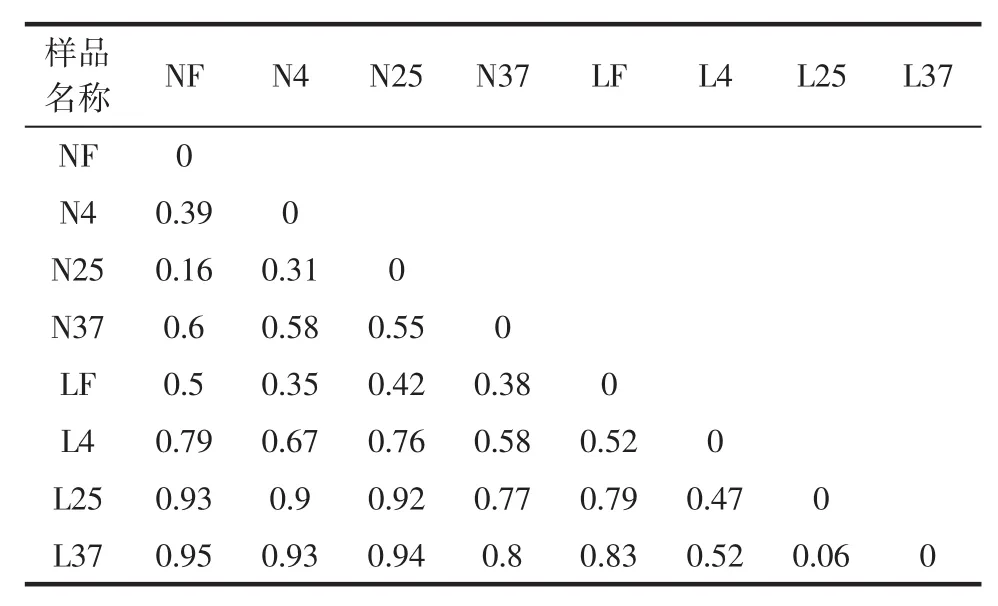
表5 样品间的距离矩阵Table 5 The distance matrix between samples
由表5可知,不同类型的豆腐的细菌多样性差异总体上小于同一类型的豆腐。内酯豆腐在新鲜状态下的物种多样性与3个温度下储藏后的变化相比于简易包装老豆腐较小,其 Beta值分别为(0.39、0.16、0.6)和(0.52、0.79、0.83),同时也表明,储藏温度越高,老豆腐的细菌多样性变化也越大,而内酯豆腐没有呈现此规律,在25℃储藏下相比于4℃与新鲜的豆腐更为相似。其中,样品L37和L25最为相似,Beta值只有0.06,除此之外,L37与其他所有样品的差异均较大,Beta值都超过了0.5。
3 结论
本研究利用MiSeq高通量测序方法分析了8个豆腐样品的腐败菌菌相,基于该技术对不同温度储藏腐败后的盒装内酯豆腐和简易包装老豆腐中菌相进行了比较,结论如下:引起内酯豆腐腐败变质的主要细菌是耐热的芽孢杆菌属,而对于简易包装的老豆腐,由于生产工艺不同,包装方式不同,在加工及运输过程中容易被污染,因此它的初始菌相构成更加复杂。老豆腐在不同温度储藏腐败后,与初始菌相差异明显,其中某些种类的细菌更容易在此条件下繁殖代谢,最终变成优势腐败菌,而其它种类细菌在后期相互竞争中减少或者消亡。4℃下老豆腐腐败菌主要有假单胞菌,气单胞菌,芽孢杆菌,25、37℃储藏腐败后明串珠菌属在数量上占据绝对优势。在复杂的食品体系中,微生物之间会通过互惠共生、群体感应或者拮抗作用等相互影响,腐败菌代谢会产粘液、产酸、产腐败味等,导致颜色、质构、气味等显现出腐败特性。如果某种细菌具有较强竞争力并且能适应此种环境,它就会大量生长繁殖成为优势菌,其他菌生长受到抑制。
本文仅分析了不同温度储藏腐败后细菌菌相的组成情况,而不同的微生物具体在豆腐的腐败过程中如何起作用需要进一步探索,这对控制豆腐中腐败菌生长,延长豆腐的保质期有着重要意义。
[1] 谷大海,常青,刘华戎.豆腐的研究概况与发展前景[J].农产品加工·创新版,2009(6):76-78
[2] 籍保平,李博.豆制品安全生产与品质控制[M].北京:化学工业出版社,2005
[3] 佟婷婷,田丰伟,王刚,等.基于宏基因组分析四川泡菜母水作引子的泡菜发酵过程中细菌多样性变化[J].食品工业科技,2015,36(21):173-177
[4] Rinke C,Schwientek P,Sczyrba A,et al.Insights into the phylogeny and coding potential of microbial dark matter[J].Nature,2013,499(7459):431-437
[5] 夏围围,贾仲君.高通量测序和DGGE分析土壤微生物群落的技术评价[J].微生物学报,2014,54(12):1489-1499
[6] Kurokawa K,Itoh T,Kuwahara T,et al.Comparative Metagenomics Revealed Commonly Enriched Gene Sets in Human Gut Microbiomes[J].DNA Research,2007,14(4):169-181
[7] Zhang X,Yue S,Zhong H,et al.A diverse bacterial community in an anoxic quinoline-degrading bioreactor determined by using pyrosequencing and clone library analysis[J].Applied Microbiology and Biotechnology,2011,91(2):425-434
[8] 向文良,罗海,梁华忠,等.基于16S-23S rRNA ITS AFLP对米酒发酵过程中原核微生物的演替分析[J].酿酒科技,2010,2010(2):43-46
[9] Połka J,Rebecchi A,Pisacane V,et al.Bacterial diversity in typical Italian salami at different ripening stages as revealed by highthroughput sequencing of 16S rRNA amplicons[J].Food Microbiology,2015,46:342-356
[10]薛静.不同贮藏温度下即食生制章鱼品质变化及其菌相分析[D].杭州:浙江工商大学,2016
[11]邓勇,滕刚.盒装豆腐货架期延长的研究[C].全国大豆蛋白加工利用及新产品开发研讨会论文汇编,1998
[12]李博.GDL豆腐中的主要腐败菌的研究及HACCP的建立[D].北京:中国农业大学,2001
[13]綦国红,董明盛,陈晓红,等.食源假单胞菌16S rRNA鉴定及其食品腐败特性的研究[J].食品工业科技,2006(11):49-51
[14]Połka J,Rebecchi A,Pisacane V,et al.Bacterial diversity in typical Italian salami at different ripening stages as revealed by highthroughput sequencing of 16S rRNA amplicons[J].Food Microbiology,2015,46 342-356
[15]李除夕,董明盛,陈晓红,等.豆腐中库特氏菌生长动力学模型和货架期预测[J].农业工程学报,2009,25(S1):82-86
The Comparison of Spoilage Microflora of Tofu at Different Storage Temperatures
WU Li-ying,CHENG Yu-liang,GUO Ya-hui,SUN Xiu-lan,QIAN He*
(The School of Food Science and Technology,Jiangnan University,Wuxi 214122,Jiangsu,China)
Studied with boxed lactone tofu and simply packaged firm tofu,including these two kinds of fresh tofus and their spoiled tofu stored at different levels of temperatures(4,25,37 ℃)respectively.Samples were collected for genome extraction and the V3 parts of 16S rRNA of each sample's genome were amplified by PCR.MiSeq high-throughput sequencing technologies was used to analyze the spoilage microflora and bacterial diversity of these two kinds of tofu.The results showed that,the microflora in boxed lactone tofu was less complicated than that in simply packaged firm tofu.The dominant bacteria were Bacillus in boxed lactone tofu and the abundance were all above 90%.In addition,the microbial diversity of the spoiled simply packaged firm tofu stored at 4℃was relatively higher compared to those in the other two simply packaged firm tofu samples.Leuconostoc was the dominant bacteria in spoiled simply packaged firm tofu stored at 25℃and 37℃,accounting for 71%and 80%respectively.Compared to the traditional culture techniques,high-throughput sequencing technologies can reflect bioinformatics which is more closed to the real status of samples.This research can provide some reference about sanitation control for small tofu production enterprises.
boxed lactone tofu;simply packaged firm tofu;high-throughput sequencing;spoilage microflora
2016-12-22
10.3969/j.issn.1005-6521.2017.18.037
“十三五”国家重点研发计划项目(2016YFD0401204)
吴丽樱(1991—),女(汉),硕士,研究方向:食品安全与质量控制。
*通信作者
猜你喜欢
读报参考(2024年10期)2024-04-06 12:09:54
名家名作(2021年11期)2021-11-12 15:16:22
艺术大观(2019年15期)2019-09-28 07:12:24
食用菌(2019年1期)2019-02-11 19:11:13
经济技术协作信息(2018年22期)2019-01-19 03:00:06
环球时报(2017-07-04)2017-07-04 08:20:21
进出口经理人(2016年11期)2016-12-17 23:15:19
科学启蒙(2016年11期)2016-12-09 19:06:45
山东青年(2016年3期)2016-02-28 14:25:59
作文评点报·小学三、四年级(2015年30期)2016-01-07 23:46:16
