
α-平滑肌肌动蛋白对高糖诱导的肾小管上皮细胞增殖、凋亡的影响
2017-09-03 10:27索艳谷优优王肃
山东医药 2017年30期
索艳,谷优优,王肃
(天津市第五中心医院,天津300451)
α-平滑肌肌动蛋白对高糖诱导的肾小管上皮细胞增殖、凋亡的影响
索艳,谷优优,王肃
(天津市第五中心医院,天津300451)
目的 探讨α-平滑肌肌动蛋白(α-SMA)对高糖诱导的肾小管上皮细胞增殖、凋亡的影响及机制。方法 培养人肾小管上皮细胞,分为低糖组、高糖组和干扰组。干扰组将α-SMA siRNA转染至肾小管上皮细胞中,高糖组、低糖组细胞均不进行α-SMA转染。转染完成后,高糖组、干扰组细胞用高糖DMEM细胞培养液培养,低糖组细胞用低糖DMEM细胞培养液培养,均培养48 h。采用MTT法检测细胞增殖情况。采用流式细胞术检测细胞凋亡情况,计算细胞凋亡率。采用Western blotting法检测细胞含半胱氨酸的天冬氨酸蛋白水解酶3(Caspase-3)、Bcl-2相关X蛋白(Bax)、Toll样受体4(TLR4)、核因子-κB(NF-κB) 蛋白表达,采用二氯二氢荧光素-乙酰乙酸酯(DCFH-DA)法检测细胞中ROS水平,采用ELISA法检测上清液TNF-α、IL-8水平。结果 高糖组、干扰组细胞增殖能力均低于低糖组(P均<0.01),干扰组细胞增殖能力高于高糖组(P<0.01)。高糖组、干扰组细胞凋亡率均高于低糖组(P均<0.01),干扰组细胞凋亡率低于高糖组(P<0.01)。高糖组、干扰组细胞Caspase-3、Bax、TLR4、NF-κB蛋白表达均高于低糖组(P均<0.01),干扰组细胞Caspase-3、Bax、TLR4、NF-κB蛋白表达均低于高糖组(P均<0.01)。高糖组、干扰组细胞中ROS水平及上清液TNF-α、IL-8水平均高于低糖组(P均<0.01),干扰组细胞中ROS水平及上清液TNF-α、IL-8水平均低于高糖组(P均<0.01)。结论 干扰α-SMA后能够逆转高糖对肾小管上皮细胞的增殖抑制和凋亡促进作用,其机制可能与降低细胞内ROS水平及抑制TLR4/NF-κB信号通路有关。
糖尿病肾病;α-平滑肌肌动蛋白;细胞增殖;细胞凋亡;炎症因子
糖尿病肾病是由糖尿病引发的慢性疾病,是终末期肾病的主要原因之一。肾小管上皮萎缩及肾组织纤维化是糖尿病肾病的主要病理变化,而肾小管上皮细胞的增殖、凋亡是导致肾小管上皮萎缩的重要原因[1]。α-平滑肌肌动蛋白(α-SMA)是肾组织纤维化的重要标志蛋白[2]。研究表明,糖尿病肾病患者肾组织中α-SMA表达增多,肾小管上皮细胞凋亡增多[3]。肾组织炎症也是糖尿病损害肾组织的重要原因。研究发现,肾组织中炎症反应与Toll样受体4(TLR4)有关,TLR4激活后可导致核因子-κB(NF-κB)活化,诱导炎症反应的发生,进而影响细胞的生长及凋亡[4]。2015年12月~2016年11月,我们采用小干扰RNA(siRNA)技术抑制人肾小管上皮细胞α-SMA蛋白表达,观察对高糖环境下肾小管上皮细胞增殖、凋亡的影响并探讨其机制,为明确α-SMA对肾小管上皮细胞增殖、凋亡影响的机制提供实验依据。
1 材料与方法
1.1 细胞及试剂 人肾小管上皮细胞HK-2购自上海中乔新舟生物科技有限公司。α-SMA多克隆抗体、活化的含半胱氨酸的天冬氨酸蛋白水解酶3(Caspase-3)单克隆抗体、Bcl-2相关X蛋白(Bax)单抗、TLR4多抗、NF-κB多抗、甘油醛-3-磷酸脱氢酶(GAPDH)单抗均购自美国Abcam公司;低糖DMEM培养基(含5.6 mmol/L的D-葡萄糖)、高糖DMEM培养基(含25 mmol/L的D-葡萄糖)均购自美国Gibco公司;活性氧(ROS)检测试剂盒、二喹啉甲酸(BCA)蛋白浓度检测试剂盒购自碧云天生物技术研究所;TNF-α、IL-8 ELISA检测试剂盒均购自上海心语生物科技有限公司。
1.2 细胞培养 液氮中保存的肾小管上皮细胞在37 ℃融化,用含有10%胎牛血清的低糖DMEM培养液悬浮细胞,接种到细胞瓶中,37 ℃、饱和湿度、5%CO2培养箱培养。待细胞融合度达到90%时,用0.25%的胰蛋白酶消化,按照1∶3比例接种至细胞瓶中继续培养。
1.3 细胞分组、转染及干预方法 取培养至对数生长期的人肾小管上皮细胞,胰蛋白酶消化后,以2×105个/孔细胞接种至6孔细胞培养板中,用不含胎牛血清的低糖培养液同步化24 h后,分为低糖组、高糖组和干扰组。干扰组按照Lipofectamine 2000转染试剂说明书将α-SMA siRNA转染至肾小管上皮细胞中,α-SMA siRNA 正义链为5′-GAGGAUGAACUGUUUCAAGTT-3′,反义链为5′-CUUGAAACAGUUCAUCCUCTT-3′。并设立对照细胞。Western blotting法检测结果示干扰组细胞α-SMA蛋白表达明显低于对照细胞,表明α-SMA siRNA转染成功。高糖组、低糖组细胞均不进行α-SMA转染。待转染完成后,高糖组、干扰组细胞用D-葡萄糖浓度为25 mmol/L的高糖DMEM细胞培养液培养,低糖组细胞用D-葡萄糖浓度为5.6 mmol/L的低糖DMEM细胞培养液培养,均培养48 h。
1.4 细胞增殖情况观察 采用MTT法。取各组细胞,每组设置6个复孔,在每孔中加入5 mg/mL的MTT溶液20 μL,放在37 ℃孵育4 h后,弃上清液,加入二甲基亚砜150 μL,摇床震荡反应10 min。酶标仪检测490 nm处的光密度(OD)值,分析细胞增殖情况。
1.5 细胞凋亡情况观察 采用流式细胞术。取各组细胞,加入胰蛋白酶消化细胞,弃上清液,用PBS悬浮细胞,调整细胞密度为1×106/mL。吸取1 mL细胞悬浮液,离心,加入500 μL的结合缓冲液混合后,加入碘化丙啶(PI)和膜联蛋白-V各5 μL,放置于室温条件下反应10 min,流式细胞仪检测细胞凋亡情况,计算细胞凋亡率。
1.6 细胞 Caspase-3、Bax、TLR4、NF-κB 蛋白表达检测 采用Western blotting法。取各组细胞,提取细胞总蛋白,Western blotting法检测细胞中 Caspase-3、Bax、TLR4、NF-κB蛋白表达。
1.7 细胞中ROS水平检测 采用二氯二氢荧光素-乙酰乙酸酯(DCFH-DA)荧光探针法。取各组细胞,胰蛋白酶消化,收集1×106个细胞,用PBS反复洗涤后,加入2′7′-DCFH-DA,放在37 ℃孵育30 min。PBS洗涤3次,荧光酶标仪检测荧光强度,检测的发射波长为530 nm,激发波长为485 nm,荧光强度越强代表ROS水平越高。
1.8 细胞上清液TNF-α、IL-8水平检测 采用ELISA法。取各组细胞,吸取培养液上清,按照ELISA检测试剂盒说明书检测培养液上清中TNF-α、IL-8水平。
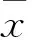
2 结果
2.1 各组细胞增殖情况比较 低糖组、高糖组、干扰组OD值分别为0.63±0.06、0.34±0.04、0.49±0.04。高糖组、干扰组细胞增殖能力均低于低糖组(P均<0.01),干扰组细胞增殖能力高于高糖组(P<0.01)。
2.2 各组细胞凋亡率比较 低糖组、高糖组、干扰组细胞凋亡率分别为11.32%±1.02%、39.25%±2.12%、19.36%±1.85%。高糖组、干扰组细胞凋亡率均高于低糖组(P均<0.01),干扰组细胞凋亡率低于高糖组(P<0.01)。
2.3 各组细胞 Caspase-3、Bax、TLR4、NF-κB 蛋白表达比较 高糖组、干扰组细胞中 Caspase-3、Bax、TLR4、NF-κB蛋白表达均高于低糖组(P均<0.01),干扰组细胞中 Caspase-3、Bax、TLR4、NF-κB蛋白表达均低于高糖组(P均<0.01)。见表1。
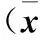
表1 各组细胞 Caspase-3、Bax、TLR4、NF-κB蛋白表达比较±s)
注:与低糖组比较,*P<0.01;与高糖组比较,#P<0.01。
2.4 各组细胞中ROS及上清液TNF-α、IL-8水平比较 高糖组、干扰组细胞中ROS水平及上清中TNF-α、IL-8水平均高于低糖组(P均<0.01),干扰组细胞中ROS水平及上清中TNF-α、IL-8水平均低于高糖组(P均<0.01)。见表2。
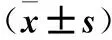
表2 各组细胞中ROS及上清液TNF-α、IL-8水平比较
注:与低糖组比较,*P<0.01;与高糖组比较,#P<0.01。
3 讨论
糖尿病肾病是一种常见的发生于肾组织的慢性疾病。肾小管上皮细胞凋亡及肾组织中的炎症是导致肾组织损伤的重要途径[5]。α-SMA是肌动蛋白家族的成员之一,在糖尿病肾病肾组织损伤中发挥重要作用[6]。研究表明,在糖尿病肾病患者肾组织中α-SMA表达增多,与肾组织的损伤程度有关[7]。目前对于α-SMA在糖尿病发病中的作用机制尚不明确,对其在高糖环境下对肾小管上皮细胞增殖、凋亡的影响及机制尚不明确。
肾小管上皮细胞的增殖、凋亡与糖尿病肾病肾组织损伤密切相关,而细胞凋亡受到一系列基因的严格调控[8]。α-SMA能够影响成纤维细胞、肾小管上皮细胞等细胞凋亡过程[9]。本研究发现,高糖组、干扰组细胞增殖能力均低于低糖组,干扰组细胞增殖能力高于高糖组,提示高糖对肾小管上皮细胞的增殖具有抑制作用,抑制α-SMA表达后,则可逆转高糖对肾小管上皮细胞的增殖抑制作用;高糖组细胞凋亡率均高于低糖组,干扰组细胞凋亡率低于高糖组,提示高糖可明显促进肾小管上皮细胞的凋亡,抑制α-SMA表达后,则可逆转高糖对肾小管上皮细胞的促凋亡作用。以上结果表明,高糖能够诱导肾小管上皮细胞凋亡、抑制肾小管上皮细胞增殖,干扰α-SMA表达后可使高糖环境下的肾小管上皮细胞增殖能力降低、凋亡增加。
Caspase-3和Bcl-2在细胞凋亡过程中发挥促进作用[10]。Bax是Bcl-2蛋白家族的成员之一,其表达升高后能够促进细胞凋亡的发生,是一种促凋亡蛋白。细胞内的ROS水平与细胞凋亡也有密切关系[11]。研究[12,13]表明,在糖尿病肾病肾组织中肾小管上皮细胞凋亡增多,ROS水平增高,促凋亡蛋白Bax表达增多,Caspase-3活化水平升高。α-SMA一方面能够提高细胞中ROS水平,使细胞内的脂质发生过氧化,造成细胞膜通透性改变,影响细胞内外平衡;另一方面可以促进细胞中凋亡相关蛋白的表达,激活细胞中Caspase级联反应,使Caspase活化成为 Caspase-3,而 Caspase-3是细胞凋亡进入不可逆阶段的标志,从而发挥促进细胞凋亡的作用;同时还能够促进Bax的表达,进一步促进细胞凋亡[14]。本研究发现,高糖组细胞中 Caspase-3、Bax蛋白及ROS表达水平均高于低糖组,提示高糖能够通过影响细胞中凋亡相关蛋白及ROS水平促进肾小管上皮细胞凋亡;干扰组细胞中 Caspase-3、Bax蛋白及ROS表达水平均低于高糖组,提示抑制α-SMA表达后,能够使高糖诱导的肾小管上皮细胞中 Caspase-3、Bax蛋白表达降低,减轻氧化应激反应,可能是其拮抗高糖对肾小管上皮细胞的增殖抑制作用、促凋亡的机制之一。
糖尿病肾病本质上也是肾组织中的炎症反应。研究发现,糖尿病肾病患者的肾组织中TNF-α、IL-8表达上升[15,16]。本研究发现,高糖组细胞上清液TNF-α、IL-8水平均高于低糖组,干扰组细胞上清液TNF-α、IL-8水平均低于高糖组,提示高糖能够诱导肾小管上皮细胞分泌TNF-α、IL-8导致炎症反应,而抑制α-SMA表达后能够拮抗高糖诱导的肾小管上皮细胞分泌TNF-α、IL-8。
Toll样受体是一种跨膜蛋白,能够将细胞外的抗原信息传递至细胞内,在免疫反应中发挥重要作用[17,18]。在肾组织中,Toll除了参与免疫反应外,还能够将信号传递给NF-κB,而NF-κB在细胞内多种转录过程中都发挥关键作用,进而影响细胞的生长[19]。TLR4/NF-κB信号转导通路在糖尿病肾病中发挥重要作用[20]。研究表明,在糖尿病肾病大鼠肾组织中发现TLR4和NF-κB的异常升高,高糖环境能够诱导大鼠肾小管上皮细胞TLR4和NF-κB表达升高[21,22]。本研究发现,高糖组细胞中TLR4、NF-κB蛋白表达均高于低糖组,干扰组细胞中TLR4、NF-κB蛋白表达均低于高糖组,这提示高糖能够诱导肾小管上皮细胞中TLR4和NF-κB升高,而抑制α-SMA表达后,肾小管上皮细胞中TLR4和NF-κB表达降低,表明抑制α-SMA表达能够拮抗高糖诱导的肾小管上皮细胞过表达TLR4和NF-κB。
综上所述,高糖可抑制肾小管上皮细胞增殖,诱导肾小管上皮细胞凋亡;抑制α-SMA表达能够拮抗高糖对肾小管上皮细胞增殖的抑制作用及促凋亡作用,其机制可能与抑制TLR4/NF-κB信号通路及降低细胞内ROS水平有关。
[1] Kitada M, Ogura Y, Suzuki T, et al. A very-low-protein diet ameliorates advanced diabetic nephropathy through autophagy induction by suppression of the mTORC1 pathway in Wistar fatty rats, an animal model of type 2 diabetes and obesity[J]. Diabetologia, 2016, 59(6):1307-1317.
[2] Chen N, Hao J, Li L, et al. Carboxy-terminal modulator protein attenuated extracellular matrix deposit by inhibiting phospho-Akt, TGF-β1 and α-SMA in kidneys of diabetic mice[J]. Biochem Biophys Res Commun, 2016,474(4):753-760.
[3] Takamiya Y, Fukami K, Yamagishi S, et al. Experimental diabetic nephropathy is accelerated in matrix metalloproteinase-2 knockout mice[J]. Nephrol Dial Transplant, 2013,28(1):55-62.
[4] Ye EA, Steinle JJ. MiR-146a protects primary human retinal microvascular endothelial cells from hyperglycemia-induced apoptosis by attenuating inflammatory pathways mediated by TLR4/NF-κB and TNF alpha[J]. Mediators Inflamm, 2016,57(12):5432.
[5] Sallustio F, Costantino V, Cox SN, et al. Human renal stem/progenitor cells repair tubular epithelial cell injury through TLR2-driven inhibin-A and microvesicle-shuttled decorin[J]. Kidney Int, 2013,83(3):392-403.
[6] 张琳琪,吕雁,刘红亮,等.益肾化瘀方对单侧输尿管结扎大鼠肾组织中 α-SMA、E-cadherin 表达的影响[J].中华中医药杂志, 2014,29(6):1966-1969.
[7] He T, Xiong J, Nie L, et al. Resveratrol inhibits renal interstitial fibrosis in diabetic nephropathy by regulating AMPK/NOX4/ROS pathway[J]. J Mol Med (Berl), 2016,94(12):1359-1371.
[8] Kang HM, Ahn SH, Choi P, et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development[J]. Nat Med, 2015,21(1):37-46.
[9] 黄凌虹,张国强.细胞增殖与凋亡在单侧输尿管梗阻大鼠肾间质纤维化发病中的意义[J].中华肾脏病杂志,2000,16(1):24-27.
[10] 姜爱英,孙迎燕,王松,等.老年晚期非小细胞肺癌组织 p53、bcl-2、caspase-3表达变化及其与预后关系[J].中国老年学杂志, 2016, 36(15): 3735-3736.
[11] Ralph SJ, Pritchard R, Rodríguez ES, et al. Hitting the bull′s-eye in metastatic cancers-NSAIDs elevate ROS in mitochondria, inducing malignant cell death[J]. Pharmaceuticals, 2015,8(1):62-106.
[12] Andeen NK, Nguyen TQ, Steegh F, et al. The phenotypes of podocytes and parietal epithelial cells may overlap in diabetic nephropathy[J]. Kidney Int, 2015,88(5):1099-1107.
[13] Wang H, Zhang H, Chen X, et al. The decreased expression of electron transfer flavoprotein is associated with tubular cell apoptosis in diabetic nephropathy[J]. Int J Mol Med, 2016,37(5):1290-1298.
[14] Li Q, Liu T, Zhang L, et al. The role of bFGF in down-regulating α-SMA expression of chondrogenically induced BMSCs and preventing the shrinkage of BMSC engineered cartilage[J]. Biomaterials, 2011,32(21):4773-4781.
[15] Sun L, Kanwar YS. Relevance of TNF-α in the context of other inflammatory cytokines in the progression of diabetic nephropathy[J]. Kidney Int, 2015,88(4):662-665.
[16] Barutta F, Bruno G, Grimaldi S, et al. Inflammation in diabetic nephropathy: moving toward clinical biomarkers and targets for treatment[J]. Endocrine, 2015,48(3):730-742.
[17] Zikou X, Tellis CC, Rousouli K, et al. Differential membrane expression of toll-like receptors and intracellular cytokine induction in peripheral blood monocytes of patients with chronic kidney disease and diabetic nephropathy[J]. Nephron Clin Pract, 2015,128(3-4):399-406.
[18] Lv J, Chen Q, Shao Y, et al. Cross-talk between angiotensin-Ⅱ and toll-like receptor 4 triggers a synergetic inflammatory response in rat mesangial cells under high glucose conditions[J]. Biochem Biophys Res Commun, 2015,459(2):264-269.
[19] Lazaro I, Oguiza A, Recio C, et al. Targeting HSP90 ameliorates nephropathy and atherosclerosis through suppression of NF-κB and STAT signaling pathways in diabetic mice[J]. Diabetes, 2015, 64(10):3600-3613.
[20] Ni H, Jin W, Zhu T, et al. Curcumin modulates TLR4/NF-κB inflammatory signaling pathway following traumatic spinal cord injury in rats[J]. J Spinal Cord Med, 2015, 38(2):199-206.
[21] Wang Y, Nie M, Lu Y, et al. Fucoidan exerts protective effects against diabetic nephropathy related to spontaneous diabetes through the NF-κB signaling pathway in vivo and in vitro[J]. Int J Mol Med, 2015,35(4):1067-1073.
[22] Wang YL, Wang K, Yu SJ, et al. Association of the TLR4 signaling pathway in the retina of streptozotocin-induced diabetic rats[J]. Graefes Arch Clin Exp Ophthalmol, 2015,253(3):389-398.
Effects of α -SMA on proliferation and apoptosis of renal tubular epithelial cells induced by high glucose
SUOYan,GUYouyou,WAMGSu
(TianjinFifthCentralHospital,Tianjin300451,China)
Objective To investigate the effects and mechanism of α-smooth muscle actin (α-SMA) on the proliferation and apoptosis of renal tubular epithelial cells induced by high glucose. Methods We cultured the human renal tubular epithelial cells and divided them into the low glucose group, high glucose group, and interference group. In the interference group, we transfected the α-SMA siRNA into the renal tubular epithelial cells. After transfection, the high glucose group and the interference group were cultured in the high glucose DMEM cell culture medium with D-glucose concentration of 25 mmol/L, the low sugar group was cultured in the low glucose DMEM cell culture medium with D-glucose concentration of 5.6 mmol/L, and they were all cultured for 48 h. MTT assay was used to detect cell proliferation. The apoptosis was detected by flow cytometry and apoptosis rate was calculated. Western blotting was used to detected the Caspase-3, Bcl-2-related X protein (Bax) and Toll-like receptor 4 (TLR4) and nuclear factor (NF)-κB protein expression. The DCFH-DA was used to detected ROS level. The levels of TNF-α and IL-8 in the supernatant were detected by ELISA.Results The cell proliferation in the high glucose group and the interference group were lower than that in the low sugar group (P<0.01), the ability of cell proliferation in the interference group was higher than that in the high glucose group (P<0.01). The apoptosis rate in the high glucose group and the interference group was higher than that in the low sugar group (P<0.01), and the rate of apoptosis in the interference group was lower than that in the high glucose group (P<0.01). The expression of Caspase-3, Bax, TLR4, and NF- κB protein in the high glucose group and the interference group was higher than that in the low sugar group (allP<0.01), the expressions of Caspase-3, Bax, TLR4, and NF- κ B protein in the interference group was lower than that in the high glucose group (allP<0.01). The levels of ROS and TNF-α, IL-8 in the supernatant in the high glucose group and the interference group were higher than those in the low sugar group (allP<0.01), the levels of ROS, and TNF-α, IL-8 in the supernatant of the interference group were lower than those in the high glucose group (allP<0.01). Conclusion Interfering with α-SMA can partially reverse the effect of high glucose on the proliferation and apoptosis of renal tubular epithelial cells by decreasing the intracellular ROS level and inhibiting the TLR4/NF- κB signaling pathway.
diabetic nephropathy; α-smooth muscle actin; cell proliferation; cell apoptosis; inflammation
10.3969/j.issn.1002-266X.2017.30.004
R692.6;R587.1
A
1002-266X(2017)30-0014-04
2017-03-10)
猜你喜欢
日用电器(2022年7期)2022-09-07
中国果业信息(2021年7期)2021-12-01
凤凰生活(2019年8期)2019-08-16
中成药(2018年6期)2018-07-11
中成药(2017年8期)2017-11-22
食品安全导刊(2016年7期)2016-05-14
中外医疗(2015年11期)2016-01-04
医学研究杂志(2015年8期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
长江蔬菜(2015年3期)2015-03-11
