
禽源H9N2亚型猪流感病毒反向遗传操作技术平台的建立
2017-06-05 15:19:57刘晓敏杨海明宫晓倩阮宝阳单同领童光志
中国动物传染病学报 2017年2期
刘晓敏,汪 琪,杨海明,宫晓倩,阮宝阳,张 鹏,单同领,童 武,童光志,于 海
·研究论文·
禽源H9N2亚型猪流感病毒反向遗传操作技术平台的建立
刘晓敏,汪 琪,杨海明,宫晓倩,阮宝阳,张 鹏,单同领,童 武,童光志,于 海
利用RT-PCR技术扩增了禽源H9N2亚型猪流感病毒A/Swine/Guangxi/7/2007(H9N2)的8个目的基因片段,分别克隆至pBD双向转录表达载体。将8个重组质粒纯化后共转染293T细胞,54 h后收集上清,接种MDCK细胞。当拯救的病毒在MDCK细胞上增殖至第3代时,收集的细胞上清经检测具有血凝效价。经过测序分析,确定获救病毒的8个基因片段序列与野生株完全一致,表明病毒拯救成功。禽源H9N2亚型猪流感病毒反向遗传操作技术平台的成功建立为探索猪流感病毒致病机理、传播机制与基因功能研究奠定了基础。
猪流感病毒;H9N2;RT-PCR;病毒拯救;反向遗传操作平台
猪流感(swine influenza,SI)是由正粘病毒科、A型流感病毒属的猪流感病毒(Swine influenza virus,SIV)引发的一种急性、传染性呼吸道疾病,一年四季均可流行,但易爆发于晚秋、冬季、早春[1],临床上主要表现为突发、高热、咳嗽、呼吸困难、反复发作、衰竭。SI一直在猪场中普遍存在且难以根除,同时容易与其他呼吸道细菌或病毒疾病,如猪呼吸与繁殖综合征、猪链球菌、猪传染性胸膜肺炎等继发或混合感染,导致病情严重化,死亡率升高,严重影响患猪健康状态与生产性能,对养猪业危害极大[2]。由于猪呼吸道上皮细胞表面同时存在禽流感和人流感病毒受体SAa-2,3-Gal和SAa-2,6-Gal,所以禽流感和人流感病毒均可感染猪,导致猪在流感病毒的种间传播过程中,扮演着“中间宿主”的角色,成为流感病毒不同毒株发生基因重组的“混合器”以及新流行毒株产生的“孵育器”,对人类健康也造成极大的潜在威胁[3,4]。
SIV是单股负链分节段的RNA病毒,按照核蛋白和基质蛋白的不同可分为A、B、C三种型。A 型SIV根据血凝素(HA)和神经氨酸酶(NA)抗原性的差异又可划分为H1N1、H1N2、H3N2、H9N2等多种不同的血清亚型。H9N2亚型流感病毒最早为1966年研究人员从火鸡中分离获得,之后在世界范围内广泛的流行与存在[5]。我国于1994年首次从广东省病鸡中分离到H9N2亚型流感病毒[6],Markwell等于1998年首次从香港猪群中分离到H9N2亚型流感病毒。文献报道显示,H9N2亚型流感病毒在国内的宿主范围正逐渐扩大,不仅能够感染家禽、野鸟,还能感染哺乳动物,如猪和人,同时它还是1997年香港流感事件中H5N1亚型流感病毒的内部基因供体[7],也是2013年可以感染人的H7N9亚型流感病毒的内部基因供体[8,9]、H10N8亚型流感病毒的内部基因供体[10]。研究表明,H9N2亚型流感病毒的遗传结构十分复杂,抗原变异十分活跃,并已获得了在哺乳动物(猪-猪)之间水平传播的能力[11]。流感病毒很有可能通过在猪体内的适应与重组,产生流感大流行毒株,因此对H9N2亚型SIV的深入研究已势在必行。
本实验室2006年从广西省感染了高致病性PRRSV的病猪中分离到4株H9N2亚型SIV,其中1株病毒命名为A/Swine/Guangxi/7/2007(H9N2)。遗传演化分析结果表明,该病毒属于禽源A/Duck/Hong Kong/Y 280/97分支[12-14]。为探讨禽源H9N2亚型SIV的分子致病机理与传播机制,本研究将其8个目的基因片段分别克隆到pBD双向转录表达载体,共转染293T细胞,成功拯救出病毒,建立了禽源H9N2亚型SIV的反向遗传操作技术平台。
1 材料与方法
1.1 毒株与载体 禽源H9N2亚型SIV A/Swine/ Guangxi/7/2007(H9N2)由本实验室分离保存;8质粒流感病毒拯救系统双向转录表达载体pBD由中国农业科学院上海兽医研究所李泽君研究员惠赠。
1.2 菌种与细胞 E.coli DH5α感受态细胞购自TIANGEN公司;293T和MDCK细胞均由本实验室保存。
1.3 实验动物 9~11日龄SPF鸡胚购自山东省实验动物中心。
1.4 主要试剂 Pfu Ultra Ⅱ Fusion HS DNA Polymerase购自Agilent Technologies公司;Gel Extraction Kit购自OMEGA Bio-Tek公司;DNA Marker(DL 2000)购自捷瑞公司;DNA PolymeraseⅠLarge(Klenow)Fragment、T4 DNA Polymerase、BspQⅠ、T4 DNA ligase购自Biolabs公司;dATP、dTTP、dGTP、dCTP购自宝生物工程(大连)有限公司;FBS、DME M(1X)、OPT I-ME M(1X)、0.25% Trypsin-EDTA(1X)购自Gibco公司;BSA购自Sigma公司;QIAprep Spin Miniprep Kit、RNeasy Mini Kit购自QIAGEN公司;SuperScriptTM Ⅲ Firststrand Synthesis System for RT-PCR、Lipofectamine 2000 Reagent购自Invitrogen公司。
1.5 引物设计与合成 对GenBank中公布的H9N2亚型流感病毒的5' 和3' 末端保守序列进行比对,参照文献[14],利用Oligo6.0软件设计合成扩增流感病毒PB2、PB1、PA、HA、NP、NA、M、NS基因的引物及流感病毒反转录通用引物Uni-12。引物均由Invitrogen公司合成。
1.6 病毒RNA的提取及目的基因片段的扩增 首先
在9~11日龄SPF鸡胚中增殖病毒,然后按照RNeasy Mini Kit说明书提取鸡胚尿囊液中的病毒总RNA,按照SuperScriptTM Ⅲ First-strand Synthesis System for RT-PCR说明书反转录合成cDNA,以此cDNA为模板,再用高保真酶Pfu Ultra Ⅱ Fusion HS DNA Polymerase扩增8个目的基因片段:PB2、PB1、PA、HA、NP、NA、M、NS。
1.7 重组质粒的构建、鉴定及纯化 将RT-PCR扩增的8个目的基因片段进行琼脂糖凝胶电泳,切取分子质量大小相符的条带,参照Gel Extraction Kit说明书分别进行胶回收。回收的产物在dCTP、dTTP存在下,用T4 DNA Polymerase进行处理。pBD载体通过BspQⅠ酶切后进行琼脂糖凝胶电泳,切下目的条带,回收产物在dATP、dGTP存在下,用DNA PolymeraseⅠLarge(Klenow)Fragment进行处理。处理过的基因片段与载体经回收后,用T4 DNA ligase于16℃过夜连接。将10μ L连接产物转化至E.coli DH5α感受态细胞中,取100μ L涂布于含有氨 青霉素抗性的LB固体培养基,37℃培养12~14 h,然后挑取单个菌落接种于含有氨 青霉素抗性的LB液体培养基,37℃培养14~16 h。经菌液PCR鉴定为阳性的样品,取适量送生工生物工程(上海)股份有限公司进行序列测定。对测序结果分析正确的菌液大量增殖,按照QIAprep Spin Miniprep Kit说明书提取重组质粒,并对质粒浓度进行精确测定。
1.8 病毒的拯救 8个重组质粒均按照1μ g计算,分别取相应体积于同一个EP管中,再加入150μ L无血清的OPTI-MEM,充分混合均匀。取20μ L Lipofectamine 2000与150μ L无血清的OPTI-MEN充分混合均匀,室温作用5 min,然后加至已混匀的质粒中,室温作用20 min。将六孔板中已培养至最佳状态的293T细胞用无血清的OPTI-MEM洗1遍,然后将上述混合物加至293T细胞,用无血清的OPTIMEM补至1 mL,轻轻混匀,于37℃、5%CO2培养箱中培养。待转染后24 h,每孔加入1 mL含8%BSA的无血清OPTI-MEN,继续培养。转染后共54 h,收集细胞上清,接种于已培养好的MDCK细胞。当MDCK细胞呈现明显病变时,收集细胞上清。
1.9 获救病毒的鉴定 对收集的细胞上清进行血凝试验,测定病毒的血凝效价,并对血凝结果为阳性的细胞上清按照RNeasy Mini Kit说明书提取病毒的总RNA,按照SuperScriptTM Ⅲ First-strand Synthesis System for RT-PCR说明书反转录合成cDNA,然后以此为模板扩增获救病毒的8个基因片段,同时对其序列进行精确测定。获救病毒的测序结果经与野生株的相应序列进行比对,确定是否一致。
2 结果
2.1 目的基因片段的RT-PCR扩增结果 提取H9N2亚型SIV野生株的总RNA,反转录合成cDNA,然后进行目的基因片段的扩增。结果显示,通过RT-PCR扩增的PB2、PB1、PA、HA、NP、NA、M、NS这8个目的基因片段的大小与理论相符(图1)。
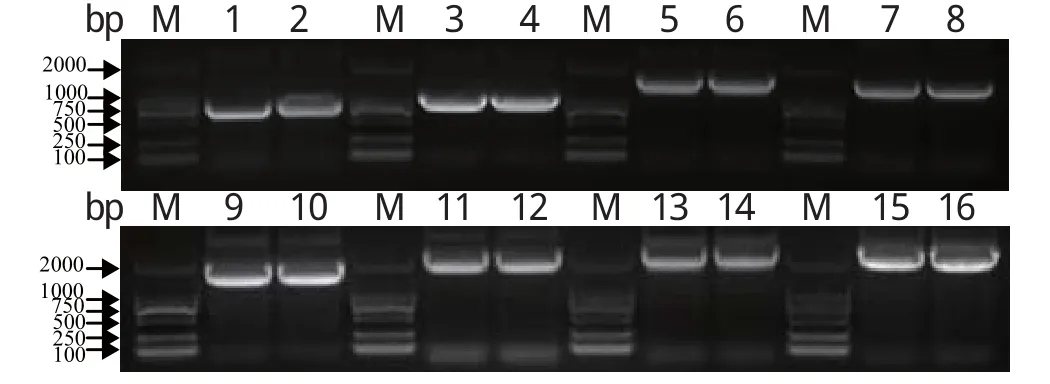
图1 H9N2亚型SIV野生株8个基因片段的RT-PCR扩增结果Fig.1 RT-PCR products of eight genes of wild type strain H9N2 SIVM: DNA分子量标准(DL2000); 1: NS基因; 2: NS基因; 3: M基因; 4: M基因; 5: NP基因; 6: NP基因; 7: NA基因; 8: NA基因; 9: HA基因; 10: HA基因; 11: PA基因; 12: PA基因; 13: PB1基因; 14: PB1基因; 15: PB2基因; 16: PB2基因M: DNA Marker (DL2000); 1: NS gene; 2: NS gene; 3: M gene; 4: M gene; 5: NP gene; 6: NP gene; 7: NA gene; 8: NA gene; 9: HA gene; 10: HA gene; 11: PA gene; 12: PA gene; 13: PB1 gene; 14: PB1 gene; 15: PB2 gene; 16: PB2 gene
2.2 重组pBD阳性质粒的鉴定结果 8个目的基因片段与双向转录表达载体pBD通过T4 DNA ligase连接后,按常规方法转化到E.coli DH5ɑ感受态细胞中,挑取单个菌落进行菌液增殖,经PCR鉴定为阳性的菌液送生工生物工程(上海)股份有限公司进行测序。
2.3 获救病毒的鉴定结果 将转染后收集的293T细胞上清接种于已培养好的MDCK细胞,待细胞呈现病变时,收集此上清。由于H9N2亚型SIV野生株在MDCK细胞增殖后的血凝效价不高,仅为21。当拯救的病毒在MDCK细胞上增殖至第3代时,对收集的细胞上清进行血凝试验,病毒效价为21,与野生株一致。将收集到的第3代细胞上清提取RNA,RTPCR扩增其8个目的基因片段,发现大小均与预期相符,PB2、PB1、PA、HA、NP、NA、M、NS分别为2341、2341、2233、1778、1565、1413、1027、890 bp(图2)。对获救病毒的8个基因片段的序列进行精确测定,结果表明获救病毒的基因序列与野生株的相应序列完全一致。由此证明病毒拯救成功,成功建立了禽源H9N2亚型SIV的反向遗传操作技术平台。
图2 rH9N2 8个基因片段的RT-PCR鉴定结果Fig.2 The identification of eight genes of rH9N2 by RTPCRM: DNA分子量标准(DL2000); 1: PB2基因; 2: PB1基因; 3: PA基因; 4: HA基因; 5: NP基因; 6: NA基因; 7: M基因; 8: NS基因M: DNA Marker (DL2000); 1: PB2 gene; 2: PB1 gene; 3: PA gene; 4: HA gene; 5: NP gene; 6: NA gene; 7: M gene; 8: NS gene
3 讨论
H9N2亚型流感病毒的宿主范围十分广泛,继Markwell等首次从猪群中分离到H9N2亚型流感病毒之后,我国研究人员陆续从猪体内分离到该病毒。2001~2003年,李海燕[15]等从猪群中分离到6株H9N2亚型流感病毒。2003年许传田等[16]从山东的发病猪群中分离到10株H9N2亚型流感病毒。2004年,动物流感重点实验室从全国的12个省中分离到41株H9N2亚型流感病毒[11]。目前研究表明,H9N2亚型流感病毒已获得在哺乳动物(猪-猪)之间水平传播的能力。1999年,郭元吉等[17]首次从广东省的流感患者体内分离到5株H9N2亚型流感病毒。Peiris等[18]从香港的儿童患者体内分离到2株H9N2亚型流感病毒。郭元吉等[17]还发现,在我国的华南和华北地区,19%的被检人群是H9N2抗体阳性。2006年包头市人群中H9亚型流感病毒抗体阳性率为36%[19]。2007年对广西壮族自治区健康青年进行的血清学调查发现,H9亚型流感病毒抗体阳性率达13.69%[20]。2008年,香港再次发生H9N2亚型流感病毒感染人的事件。这说明,H9N2亚型流感病毒比较温和,在人群中广泛存在,但不致人死亡[21],这也进一步增加了其在人群或猪群中适应与重组的可能。
按照毒力强弱划分,H9N2亚型流感病毒属于低致病性的流感病毒,在自然感染条件下,很少能够引起宿主强烈的致死反应,不过病毒各片段上氨基酸位点的改变会对病毒的毒力造成巨大影响[22],而且同一个亚型的不同毒株在致病力上也有差异[23]。研究发现,H9N2亚型流感病毒非常容易与其他亚型的流感病毒发生重组而产生新的基因型。Peiris等[18]在2001年首次证实在中国东南部地区的猪群中同时存在着H9N2和H3N2,并预测这些病毒可能通过重组而导致新病毒的产生[11]。于海等[13]对1998~2007年间从国内猪体内分离的28株H9N2亚型SIV的基因片段进行分析发现,国内猪群中至少同时存在着10种基因型(A~J),其中E~J型是H9与H5的重组病毒。有关流感专家预测,H9N2亚型流感病毒很有可能成为下次流感大爆发的元凶。因此,对SIV的分子致病机制进行深入研究显得尤为重要。
反向遗传操作技术是近年来在分子生物学研究领域非常热门的一种新型技术,是指通过构建病毒的感染性cDNA克隆,利用基因突变、缺失、插入、置换等定向修饰或改造病毒基因组序列,在共转染易感细胞或宿主的基础上重新拯救出感染性病毒粒子,再对病毒的结构和功能、复制与表达调控机理、致病机制以及病毒和宿主的相互关系等进行深入研究[24-26]。该技术的兴起和发展有助于解释由RNA病毒介导的复杂的感染过程,可促进解决RNA病毒基因组在体外难以操作的难题。目前,反向遗传操作技术技术已应用于多种负链RNA病毒的研究,特别在探究流感病毒致病机理、宿主范围限制性以及新型疫苗研制上发挥着重要作用[27,28]。本研究应用8质粒流感病毒双向转录表达系统,在cDNA水平上通过克隆对SIV进行一系列操作,继而从哺乳动物细胞中拯救出具有活性的病毒粒子,经测序验证,获救病毒的各基因片段与原始毒株相应序列完全一致。鉴于禽源H9N2亚型SIV极其重要的公共卫生学意义,其反向遗传操作平台的成功建立为接下来进一步研究病毒的种间传播、变异机制、致病性等奠定了坚实基础,也为SIV的防控和预警提供了强有力的技术支撑。
[1] Shope R E. Swine influenza III. Filtration experiments and etiology[J]. J Exp Med, 1931, 54(3): 373-385.
[2] 于海. 中国部分地区猪流感病毒的分子流行病学研究[D]. 北京: 中国农业科学院, 2008.
[3] Castrucci M R, Donatelli I, Sidoli L, et al. Genetic reassortment between avian and human influenza A viruses in Italian pigs[J]. Virology, 1993, 193(1): 503-506.
[4] Ito T, Couceiro J N, Kelm S, et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential[J]. J Virol, 1998, 72(9): 7367-7373.
[5] Homme P J, Easterday B C. Avian influenza virus infections. I. Characteristics of influenza A/Turkey/ Wisconsin/1966 virus[J]. Avian Dis, 1970, 14(1): 66-74.
[6] 陈伯伦, 张泽纪. 禽流感研究: I. 鸡A型禽流感病毒的分离与血清学初步鉴定[J]. 中国兽医杂志, 1994, 20(10): 3-5.
[7] Guan Y, Shortridge K F, K rauss S, et al. Molecular characterization of H9N2 influenza viruses: were they the donors of the “internal” genes of H5N1 viruses in Hong Kong?[J]. Proc Natl Acad Sci USA, 1999, 96(16): 9363-9367.
[8] K ageyama T, Fujisaki S, Takashita E, et al. Genetic analysis of novel avian A (H7N9) influenza viruses isolated from patients in China, February to April 2013[J]. Euro Surveill, 2013, 18(15): 7-21.
[9] Lam T T, Wang J, Shen Y, et al. The genesis and source of the H7N9 influenza viruses causing human infections in China[J]. Nature, 2013, 502(7470): 241-244.
[10] C hen H, Y uan H, Gao R, et al. C linical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: a descriptive study[J]. Lancet, 2014, 383(9918): 南宁: 714-721.
[11] 何奇松. 广西马源H9N2亚型流感病毒的分离鉴定与全基因组序列分析[D]. 南宁: 广西大学, 2012.
[12] Y u H, Hua R H, Wei T C, et al. Isolation and genetic characterization of avian origin H9N2 influenza viruses from pigs in China[J]. Vet Microbiol, 2008, 131(1): 82-92.
[13] Y u H, Zhou Y J, Li G X, et al. Genetic diversity of H9N2 influenza viruses from pigs in China: a potential threat to human health?[J]. Vet Microbiol, 2011, 149(1): 254-261. [14] Hoffmann E, Stech J, Guan Y, et al. Universal primer set for the full-length amplification of all influenza A viruses[J]. Arch Virol, 2001, 146(12): 2275-2289.
[15] 李海燕, 于康震, 杨焕良, 等. 中国猪源H5N1和H9N2亚型流感病毒的分离鉴定[J]. 中国预防兽医学报, 2004, 26(1): 1-6.
[16] 许传田, 范伟兴, 赵宏坤. A 型猪流感病毒山东分离株鉴定及其 HA 基因序列分析[J]. 中国病毒学, 2004, 19(1): 27-31.
[17] 郭元吉, 李建国, 程小雯, 等. 禽 H9N2 亚型流感病毒能感染人的发现[J]. 中华实验和临床病毒学杂志, 1999, 13(2): 105-108.
[18] Peiris M, Y uen K Y, Leung C W, et al. Human infection with influenza H9N2[J]. Lancet, 1999, 354(9182): 916-917.
[19] 张美英, 徐能义, 马淑一, 等. 包头市 2006 年人群甲型流感病毒 H1, H3, H9 抗体检测结果分析[J]. 现代预防医学, 2009, 36(20): 3958-3959.
[20] 陈妍梅, 葛万运, 黄川, 等. 广西健康青年 H9, H6 亚型禽流感病毒血清抗体调查[J]. 中国热带医学, 2008, 8(6): 985-986.
[21] 赵有淑. 3 株 H9N2 猪流感福建株的分离鉴定及分子衍化研究[D]. 甘肃: 甘肃农业大学, 2005.
[22] Zhang Z, Hu S, L i Z, et al. Multiple amino acid substitutions involved in enhanced pathogenicity of LPAI H9N2 in mice[J]. Infect Genet Evol, 2011, 11(7): 1790-1797.
[23] Gharaibeh S. Pathogenicity of an avian influenza virus serotype H9N2 in chickens[J]. Avian Dis, 2008, 52(1): 106-110.
[24] Radecke F, Billeter M A. Reverse genetics meets the nonsegmented negative-strand RNA viruses[J]. Rev Med Virol, 1997, 7(1): 49-63.
[25] Peeters B P, de Leeuw O S, Koch G, et al. Rescue of Newcastle disease virus from cloned cDNA: evidence that cleavability of the fusion protein is a major determinantfor virulence[J]. J Virol, 1999, 73(6): 5001-5009.
[26] 彭永刚, 耿庆华, 李梅. 反向遗传操作技术在流感病毒研究中的应用进展[J]. 畜牧兽医科技信息, 2007 (12): 8-10.
[27] 袁东波, 汤德元, 汪忠荣, 等. 反向遗传操作技术在甲型
流感病毒中的应用[J]. 畜牧与兽医, 2008, 40(1): 88-90. [28] Zhou B, Jerzak G, Scholes D T, et al. Reverse genetics
plasmid for cloning unstable influenza A virus gene segments[J]. J Virol Methods, 2011, 173(2): 378-383.
ESTABLISHMENT OF REVERSE GENETICS SYSTEM OF AVIAN-LIKE H9N2 SUBTYPE SWINE INFLUENZA VIRUS
LIU Xiao-min, WANG Qi, YANG Hai-ming, GONG Xiao-qian, RUAN Bao-yang, ZHANG Peng, SHAN Tong-ling, TONG Wu, TONG Guang-zhi, YU Hai
(Shanghai Veterinary Research Institute, CAAS, Shanghai 200241,China)
Eight gene fragments of avian-like H9N2 subtype Swine influenza virus A/Swine/Guangxi/7/2007(H9N2) were amplified by RT-PCR and separately cloned into the vector of pBD. Then recombinant plasmids were purified and co-transfected into 293T cells. The supernatant of 293T cells was collected after 54 h and inoculated into MDCK cells. Virus titer was determined in hemagglutination test after the rescued virus was proliferated into MDCK cells for three times, which was comparable to the wild type strain. Sequence analysis demonstrated that eight genes of the rescued virus were identical to the original virus, indicating that the virus was rescued successfully. The establishment of reverse genetics system of avian-like H9N2 subtype Swine influenza virus settles the foundation for the research on pathogenesis, transmission mechanism and gene function in the future.
Swine influenza virus; H9N2; RT-PCR; virus rescue; reverse genetics system
S852.659.5
A
1674-6422(2017)02-0029-06
2016-09-29
上海市自然科学基金青年项目(16ZR1444000);中央级公益性科研院所基本科研业务费项目(2015JB07);中国农业科学院创新工程“猪病毒性繁殖障碍综合症团队”项目
刘晓敏,女,硕士研究生,兽医学专业
童光志,E-mail: gztong@shvri.ac.cn;于海,E-mail: haiyu@shvri.ac.cn
(中国农业科学院上海兽医研究所,上海 200241)
猜你喜欢
广东医科大学学报(2020年6期)2020-02-06 06:00:40
食品科学(2018年10期)2018-05-23 01:27:28
现代检验医学杂志(2016年3期)2016-11-15 01:59:50
现代检验医学杂志(2016年1期)2016-11-12 13:19:42
分析测试学报(2015年6期)2016-01-13 06:19:03
中国病理生理杂志(2015年8期)2015-12-21 12:38:10
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年30期)2015-03-01 02:08:19
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
