
神农香菊不同株系间光合特性及叶绿素荧光特性比较分析
2017-05-19 05:34焦宏斌金利妍周蕴薇
上海农业学报 2017年2期
何 淼,刘 洋,焦宏斌,王 想,金利妍,周蕴薇
(1东北林业大学园林学院,哈尔滨 150040;2北京星河园林景观工程有限公司,北京 100018)
神农香菊不同株系间光合特性及叶绿素荧光特性比较分析
何 淼1,刘 洋1,焦宏斌2,王 想1,金利妍1,周蕴薇1
(1东北林业大学园林学院,哈尔滨 150040;2北京星河园林景观工程有限公司,北京 100018)
以成功引种栽培在东北林业大学园林学院苗圃的神农香菊(Dendranthema indicum var.aromaticum)3个优良株系S1401、S1407和S1415为研究对象,采用Li-6400XT便携式光合作用分析系统对3个株系的光合生理差异以及它们在不同光照环境的适应性进行了比较分析。结果表明:(1)S1407为双峰型曲线,S1401和S1415为单峰曲线;3个株系的光补偿点差异不大,S1415的光饱和点最高,S1401最低;对强光的利用能力S1415>S1407>S1401,S1415的光适应范围最广。(2)在荧光参数上,3个株系的初始荧光Fo大小是S1407>S1415>S1401,PhiPS2、qP、ETR均为S1401>S1407>S1415,S1401的非光化学淬灭系数qN最小,为0.83,其光合系统的自我保护能力最弱。
神农香菊;光合特性;叶绿素荧光;比较分析
神农香菊(Dendranthema indicum var.aromaticum)是野菊(Dendranthema indicum)的一个新变种[1],分布于高海拔的阳光充足地,由于当地独特的气候使其具有罕见的香气,具有平肝明目、清热解毒、散风降压的功效,具有较高的观赏和药用价值。光是影响植物生长的重要因子之一,光合作用是地球生物圈中物质和能量循环不可缺少的环节,不同种间和同一种不同品种间均存在光合特性差异,光合速率高低、光合功能期长短、光适应范围等都会影响到作物的生长状况和产量。掌握不同品种的光合作用特点和光合效能差异,有利于高光效亲本的选择和优良品种的筛选[2]。目前,国内外对神农香菊光合特性和叶绿素荧光特性方面的研究甚少,对其生物学认识不足。本研究对3个株系的光合特性方面进行比较研究,探讨了3个株系的光合能力及它们对不同光照环境的适应性进行比较分析,旨在了解同组不同株系之间光合生理的差异,为引种栽培和良种选育提供依据。
1 材料与方法
1.1 试验材料
试验材料神农香菊(Dendranthema indicum var.aromaticum)引自湖北神农架地区(110°23′57″E,31°28′7″N),在东北林业大学园林学院苗圃基地(126°37′E,45°43′N)栽培繁殖多年后,筛选出3个无性系,分别为S1401、S1407和S1415,10月下旬用细沙进行脚芽扦插,两周生根后换土,放置于温室中培养,次年2月以直径为12 cm的花盆上盆,4月移至东北林业大学园林学院112植物培养室培养,作为试验材料备用。试验中随机选取健壮的、长势一致的植株第3片成熟叶进行进行光合特性测定。
1.2 测定指标及方法
1.2.1 光合日变化的测定
选择晴朗无风天气采用美国生产的Li-6400XT光合测定仪,使用2 cm×3 cm透明叶室进行测量,选择生长健壮的植株,随机选取3株向阳面的第3片完全功能叶,在4:00—20:00每间隔2 h测定1次,共测9次,每次重复记录5个观测值,取其平均值作为该时刻的测定值。测定过程未进行温湿度和CO2的控制,气体流速控制为400μmol·s-1。读取叶片瞬时净光合速率值,同时取蒸腾速率、气孔导度、胞间二氧化碳浓度、光合有效辐射、气温、相对湿度等生理生态指标[3]。
1.2.2 光响应曲线的测定
定于晴天上午9:00—11:00采用Li-Cor公司的Li-6400XT光合测定仪进行测量,采用专用内置红蓝光源控制叶室内部光强,设定光强梯度为2 000μmol·m-2·s-1,1 800μmol·m-2·s-1,1 600μmol·m-2·s-1,1 400μmol·m-2·s-1,1 200μmol·m-2·s-1,1 000μmol·m-2·s-1,800μmol·m-2·s-1,600μmol·m-2·s-1,400μmol·m-2·s-1,200μmol·m-2·s-1,100μmol·m-2·s-1,50μmol·m-2·s-1,0μmol·m-2·s-1,设定系统内气流速度为500μmol·s-1,CO2浓度为400μmol·mol-1,温度控制在当时的外界环境温度,使用自动测量程序测定相应的净光合速率值,每组梯度重复3次。根据非直线双曲线模型公式,利用SPSS非线性回归结果,得到拟合直线方程y=ax+b,式中y为净光合速率,x为光合有效辐射,净光合速率为0时的光强为光补偿点,净光合速率最大时的光强为光饱和点。
1.2.3 叶绿素荧光诱导动力学参数的测定
采用Li-Cor公司的Li-6400光合测定仪。荧光参数设置:系统调制频率设定为0.25 kHz,闪光持续时间为0.8 s下降幅度为20%,强度为9。叶片经暗适应处理(铝箔纸包裹叶片)2 h后,测定潜在光化学效率,得到初始荧光Fo、最大荧光Fm和原初光能转化效率Fv/Fm,然后打开活化光(1 800μmol·m-2·s-1)活化1 h,再测量最小荧光Fo′、最大荧光Fm′、稳态荧光Fs和作用光存在时光反应中心实际的光化学量子效率φPSⅡ,然后根据公式:Fv/Fm=(Fm-Fo)/Fm、qP=(Fm′-Fs)/(Fm′-Fs)、qN=(Fm-Fm′)/(Fm-Fo′),计算得出光化学量子效率、光化学猝灭系数和非光化学猝灭系数值。
1.3 数据处理
试验数据采用SPSS 13.0进行统计学分析,利用Excel2003作图。
2 结果与分析
2.1 光合日变化
植物光合作用与外界环境的关系密切,一般情况下栽培植物的光合日变化呈双峰型或单峰型曲线[4]。由图1可知:3个株系神农香菊的净光合速率日变化的规律相似,S1407为双峰型曲线,S1401和S1415为单峰曲线。在上午10:00左右,S1407和S1401出现第1个峰值,净光合速率分别达到11.18μmol·m-2·s-1,10.10μmol·m-2·s-1,10:00—14:00由于光合有效辐射过强、气温升高以及空气湿度下降,净光合速率开始下降,S1401在12:00降至8.55μmol·m-2·s-1,之后基本保持不变,S1407还在14:00时跌入谷值,之后略有上升,达到第二个峰值,出现光合“午休”现象。S1415出现峰值较晚,在12:00达到峰值后净光合速率持续下降。3个株系的神农香菊16:00后净光合速率均迅速下降,19:00植物呼吸速率开始大于光合速率。
2.2 光合参数日变化
影响植物光合作用的生态因子主要包括:叶片气孔导度、胞间CO2浓度、蒸腾速率等,它们共同作用影响植物净光合速率的变化。
3个株系神农香菊的气孔导度日变化均呈双峰型(图2),与净光合速率日变化呈正相关关系。S1401和S1415的第1次峰值均出现在10:00,峰值分别为0.17mol·m-2·s-1和0.21mol·m-2·s-1,S1407的第1次峰值出现在8:00,为0.16 mol·m-2·s-1,S1401和S1407的第2次峰值均出现在16:00,S1415出现在14:00,由大至小依次为S1401(0.13 mol·m-2·s-1)>S1407(0.12 mol·m-2·s-1)>S1415(0.11 mol·m-2·s-1)。气孔导度在10:00—14:00呈下降趋势,而这期间正是光合有效辐射迅速增强,气温升高,相对湿度降低的时候,植物根据外界环境条件变化,通过控制气孔导度,调节气孔阻力,在保持正常生理活动的前提下,较少水分的散失量。
3个株系神农香菊的胞间二氧化碳浓度(图3)与净光合速率、气孔导度、蒸腾速率呈负相关关系。S1407和S1415的胞间CO2浓度均在6:00出现第1个峰值,6:00—18:00三个株系神农香菊的胞间CO2浓度持续出现低谷,说明随着白天神农香菊叶片光合作用的进行,消耗较多的CO2原料,使胞间CO2摩尔分数较低,且维持在相对稳定的水平。
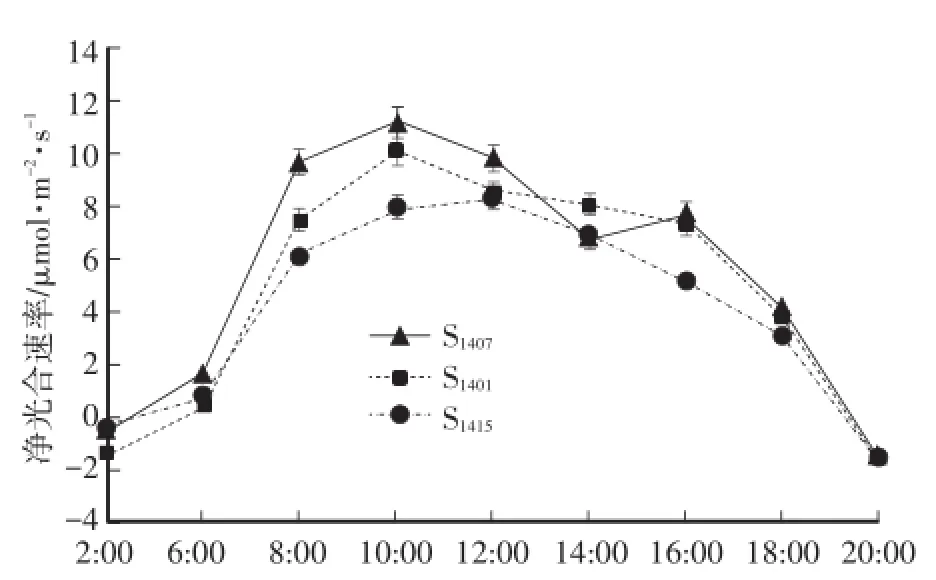
图1 3个株系神农香菊净光合速率日变化Fig.1 Diurnal variations of net photosynthetic rate of 3 lines of D.indicum var.aromaticum
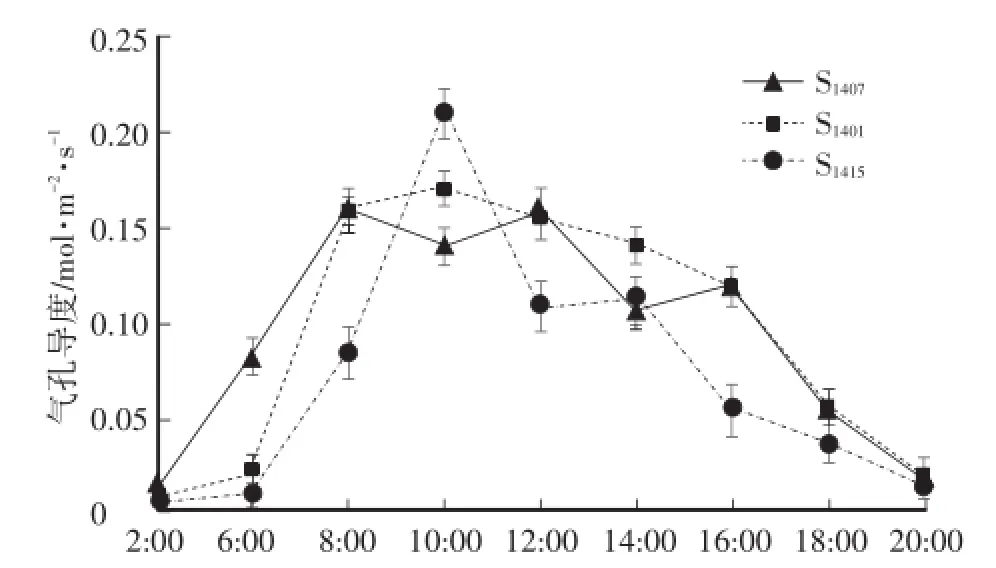
图2 3个株系神农香菊气孔导度日变化Fig.2 Diurnal variations of stomatal conductance of 3 lines of D.indicum var.aromaticum

图3 3个株系神农香菊胞间CO2浓度日变化Fig.3 Diurnal variations of intercellular CO2concentration of 3 lines of D.indicum var.aromaticum
3个株系神农香菊的蒸腾速率(图4)与净光合速率呈一定的正相关,相关系数(表1)S1401为0.94,S1407为086,S1415为0.96,也均呈现双峰曲线,出现蒸腾“午休”现象,2次出现峰值时刻分别为10:00和14:00。由图4可知:3个株系神农香菊的蒸腾速率在上午10:00都达到峰值,在下午时都呈现总体下降的趋势。S1401的蒸腾速率在10:00达到最大,为5.95 mmol·m-2·s-1,随后逐渐开始下降,18:00达到1.46 mmol·m-2·s-1,天黑后维持在0附近;S1407的蒸腾速率在10:00—12:00变化不大,在12:00达到峰值5.19 mmol·m-2·s-1后有稍微下降,然后16:00再次达到第2个峰值;S1415的蒸腾速率波动较大,在10:00时达到1个峰值为712 mmol·m-2·s-1,下降后到12:00又呈上升趋势。

图4 3个株系神农香菊蒸腾速率日变化Fig.4 Diurnal variations of transpiration rate of 3 lines of D.indicum var.aromaticum
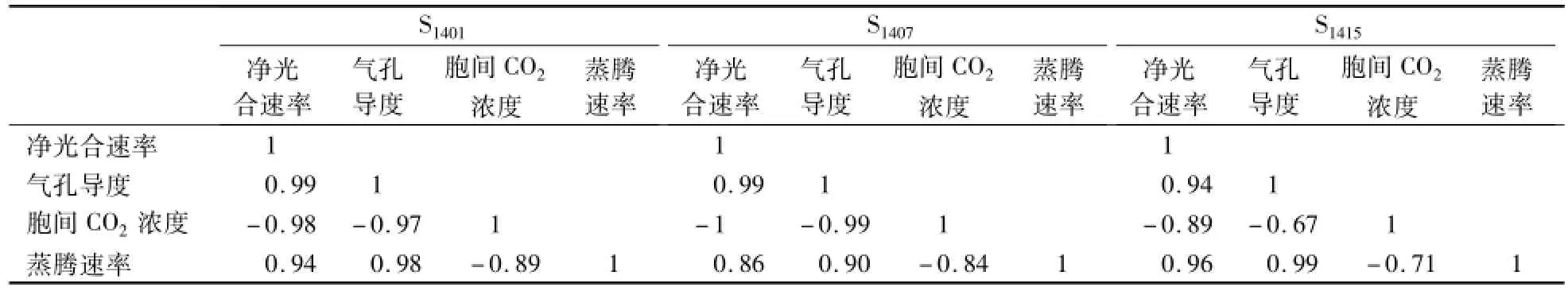
表1 3个株系神农香菊光合参数相关性分析Table 1 Correlation analysis of photosynthetic parameters of 3 lines of D.indicum var.aromaticum
2.3 光响应曲线
图5显示:3个株系神农香菊的光响应曲线变化趋势相近,在0—100μmol·m-2·s-1的光合有效辐射范围内,3个株系的净光合速率几乎呈线性增长,随着有效辐射强度的增加,净光合速率的增长速度逐渐减慢,当光合有效辐射达到1 000μmol·m-2·s-1以上时,神农香菊的净光合速率的增长缓慢,基本保持不变,叶片的光合作用处于饱和状态。
通过对比3个株系的光响应曲线,发现它们叶片的最大净光合速率差异显著,但对光合有效辐射的响应趋势基本一致,均是在低强度下迅速升高后趋于稳定。在0—100μmol·m-2·s-1时,3个株系神农香菊净光合速率变化相差不大。在光合有效辐射增大到100μmol·m-2·s-1后,3个株系间的净光合速率值出现差异,最大净光合速率差别较大,其中S1407最大,可达17.44μmol·m-2·s-1,而S1401和S1415较低。
表2表明:3个株系神农香菊的光饱和点具有一定的差异,说明它们对强光的利用能力不同,但光补偿点差异不大。在3个株系神农香菊中,S1401的光补偿点和光饱和点最低,说明其对强光的利用能力最弱,对弱光的利用能力更强;S1407具有相对较低的光补偿点,又具有较高的光饱和点和最大的净光合速率,对强光的适应能力很强,光合能力也最强;S1415的光补偿点最高,而光饱和点也最高,对光的适应范围相对最大,它的净光合速率在初期低于S1401,而当光合有效辐射超过1 600μmol·m-2·s-1时,净光合速率开始超过S1401,说明S1415利用强光的能力优于S1401。
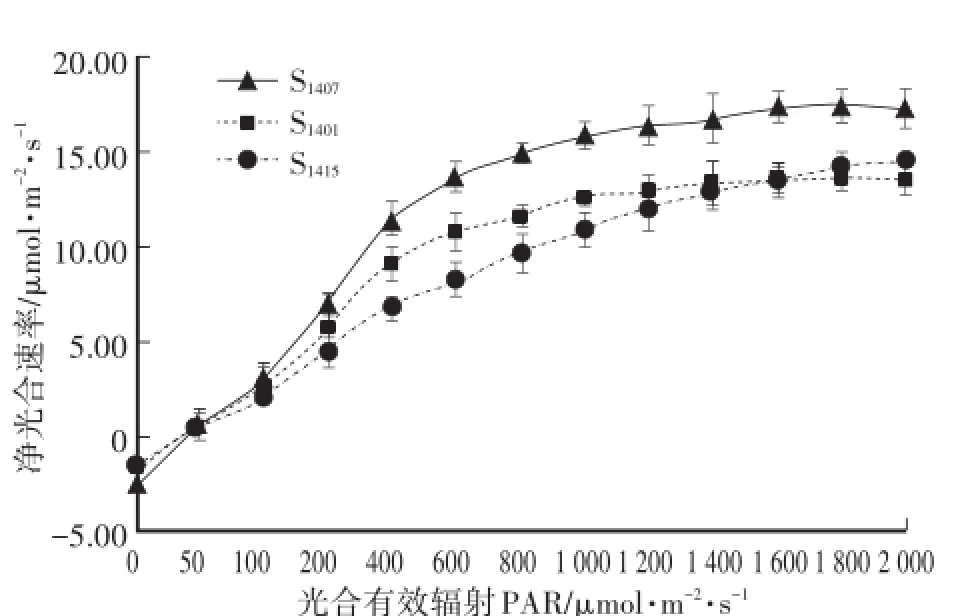
图5 3个株系神农香菊的光响应曲线Fig.5 Light response curves of 3 lines of D.indicum var.aromaticum

表2 光响应曲线方程及光饱和点和光补偿点Table 2 Light response curve equation,light saturation point and light compensation point
2.4 叶绿素荧光参数测定
2.4.1 潜在光化学效率
初始荧光Fo是PSⅡ反应中心全部开放时的荧光产量,其大小主要与天线色素到PSⅡ反应中心的激发能传递速率的结构状态及叶绿素含量有关,而与光合作用光化学反应无关[5]。Fm是暗适应下最大荧光产量,是PSⅡ反应中心处于完全关闭时的荧光产量,反映通过PSⅡ的电子传递情况[6];Fv=Fm-Fo为可变荧光,反映初级受体QA的还原情况;Fv/Fm是潜在的光化学效率,反映PSⅡ反应中心内光能转换效率,在没有环境胁迫条件下该参数都很稳定,各种植物叶片这一参数基本维持在0.80—0.85,胁迫条件下该参数明显下降。
在正常条件下,3个株系神农香菊的荧光参数如表3所示,S1407和S1415的各荧光参数差异不显著,但是与S1401差异显著。S1401的Fo和Fm分别为132.88、606.11,显著低于S1407和S1415;潜在量子效率Fv/Fm差异不显著,维持在0.8左右;Fv/Fo是Fv/Fm的另一种表达方式,它对效率的变化更加敏感,变化幅度要比Fv/Fm大的多,因此不同株系神农香菊虽然Fv/Fm差异不显著,但是S1401的Fv/Fo显著小于S1407和S1415。

表3 3个株系神农香菊潜在光化学效率Table 3 Potential photochem ical efficiencies of 3 lines of D.indicum var.aromaticum
2.4.2 PSⅡ的实际光化学效率
Fv'/Fm'为开放的PSⅡ反应中心的激发能捕获效率,可以用来估计光合机构吸收的光能被用于光化学反应和天线热耗散的相对份额;PhiPS2为作用光存在时PSⅡ实际的光化学量子效率;光化学淬灭系数qP是PSⅡ反应中心捕获的光子能量用于光化学反应的能力指标,反映了PSⅡ反应中心的开放程度;非光化学淬灭系数qN反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分,反应了植物热耗散的能力;ETR为电子传递速率,光化学淬灭系数qP愈大,初级受体QA重新氧化形成量愈大,即PSⅡ的电子传递活性愈大[7]。
在没有任何胁迫下3个株系神农香菊的实际光化学效率参数如表4所示,S1407和S1415的各荧光参数差异不显著,但是与S1401差异显著。S1401的Fv'/Fm'、PhiPS2、qP、ETR均大于S1407和S1415,其中Fv'/Fm'为S1401>S1415>S1407,而PhiPS2、qP、ETR均为S1401>S1407>S1415;S1401的非光化学淬灭系数qN最小,为0.83,显著小于S1407和S1415的0.90和0.87。

表4 3个株系神农香菊实际光化学效率和淬灭系数Table 4 Actual photochem ical efficiencies and quenching coefficients of 3 lines of D.indicum var.aromaticum
3 结论与讨论
3.1 不同株系的神农香菊光合日变化规律的差异
虽然3个株系同为神农香菊,但是在表型上存在较大差异,可能是在引种和培育过程中出现了不同方向的进化。通过分析3个株系神农香菊的光合日变化,发现神农香菊存在光合“午休”现象,且主要是受气孔限制因素影响,净光合速率的变化同时受到环境因子的影响。在相同的条件下,3个株系神农香菊的净光合速率、叶片气孔导度、胞间CO2浓度、蒸腾速率等生理因子也不完全一致,说明其光合作用除受环境因子的综合影响外,也受到自身因素的限制,同一环境因子对不同植物的影响也不同[8],与这一研究结果一致。
3.2 不同株系的神农香菊对光照强度适应性的差异
光合能力强弱(包括光饱和点和光补偿点的变化)在一定程度上具有物种遗传稳定性[7]。光补偿点较高、光饱和点较低的植物对光照的适应性较弱,反之则表明植物对光环境的适应性较强[9]。S1415对光的适应能力更强,对强光的利用率最高,而S1401适应能力最弱,但是通过荧光分析发现S1401的热耗散能力相对较好,而S1407的净光合速率最大,说明其光合能力最强,对光的利用效率也较高[10],可优先考虑引种。简在友等[11]研究表明,不同品种植物形态特征和光合生理特性之间的差异也是它们对不同生长环境的适应结果。
3.3 不同株系的神农香菊的光能转化效率的差异
S1407和S1415的各荧光参数差异不显著,但是与S1401差异显著,潜在量子效率Fv/Fm差异不显著,维持在0.8左右。表明3个株系的神农香菊未受到胁迫,且外在条件对3个株系的神农香菊的PSⅡ反应中心造成伤害[12]。肖春旺等[13]研究表明,Fo、Fm、Fv值较低,Fv/Fm和Fv/Fo值较高,说明植物的PSⅡ反应中心的光量子电子传递和转换效率较高。不同株系神农香菊虽然Fv/Fm差异不显著,但是S1401的Fv/Fo显著小于S1407和S1415且S1401的非光化学淬灭系数qN最小[14],为0.83,其光合机构的自我保护能力最弱。以上研究结果表明,不同株系的神农香菊吸收光能和热耗散能力具有较大差异。初步对比3个株系在光合特性上的差异,以期为以后神农香菊在生产栽培和植物保护的实践中提供理论[15]。
[1]杜冰群,刘启宏,朱翠英,等.两种菊属植物的核型研究[J].植物科学学报,1989,7(3):293-296.
[2]姜武.水蜜桃品种不同季节的光合特性研究[D].南京:南京农业大学,2007.
[3]金则新,柯世省.云锦杜鹃叶片光合作用日变化特征[J].植物研究,2004,24(4):447-452.
[4]高鹤,宗俊勤,陈静波,等.7种优良观赏草光合生理日变化及光响应特征研究[J].草业学报,2010,19(4):87-93.
[5]吴泽民,黄成林,韦朝领.黄山松群落林隙光能效应与黄山松的更新[J].应用生态学报,2000,11(1):13-18.
[6]GENTY B,BRIANTAIS JM,BAKER N R.The relationship between quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J].Biochim Biophys Acta,1989,990:87-92.
[7]KRAUSEG H,WEISE.Chlorophyll fluorescence and photosynthesis[J].Annu Rev Plant Physiol Plant Mol Biol,1991,42:313-349.
[8]SHARKEY T D,CHEN X,YEH S.Isoprene increases thermotolerance of fosmidomycin-fed leaves[J].Plant Physiol,2001,125:2001-2006.
[9]FARQUHAR G D.SHARKEY TD.Stomatal conductance and photosynthesis[J].Annum Rev Plant Physiol,2000,33:317-345.
[10]BIORKMAN O,HOLMGREN P.Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitat[J].Physiol Plant,1963,16(4):889-914.
[11]简在友,王文全,孟丽,等.芍药组内不同类群间光合特性及叶绿素荧光特性比较[J].植物生态学报,2010,34(12):1463-1471.
[12]惠红霞,许兴,李前荣.外源甜菜碱对盐胁迫下枸杞光合功能的改善[J].西北植物学报,2003,23(12):2137-2142.
[13]肖春旺,周广胜.毛乌素沙地中间锦鸡儿幼苗生长、气体交换和叶绿素荧光对模拟降水量变化的响应[J].应用生态学报,2001,12(5):692-696.
[14]MAXWELL K,JOHNOSN G N.Chlorophyll fluorescence-a practical guide[J].Journal of Experimental Botany,1998,51:659-668.
[15]刘周莉,赵明珠,秦娇娇,等.3种木质藤本植物光合特性比较研究[J].生态环境学报,2015,24(6):952-956.
(责任编辑:程智强)
Comparative analysis on the photosynthetic performances and chlorophyll fluorescence characteristics among lines of Dendranthema indicum var.aromaticum
HE Miao1,LIU Yang1,JIAO Hong-bin2,WANG Xiang1,JIN Li-yan1,ZHOU Yun-wei1
(1College of Landscape Architecture,Northeast Forestry University,Harbin 150040,China;2Beijing Star-River Landscape Engineering Company Limited,Beijing 100018,China)
Three superior lines of Dendranthema indicum var.aromaticum(S1401,S1407and S1415)successfully introduced and cultivated in a nursery of this college were used as testmaterials,and their difference in photosynthetic performance and adaptability to different light environments were comparatively analyzed by using the portable Li-6400 XT photosynthesismeasuring system.The results showed that(1)in terms of diurnal variation of photosynthetic rate,S1407presented a double-hump curve and S1401and S1415a unimodal curve;The 3 lines’difference was less in light compensation point,and in light saturation point S1451was the highest and S1401the lowest;The descending order of ability to utilize strong lightwas S1415>S1407>S1401,and S1415had the widest light adaptation range.(2)In terms of fluorescence parameters,the order of 3 lines was S1407>S1415>S1401in initial fluorescence Fo values and S1401>S1407>S1415in PhiPS2,qP and ETR values,and the non-photochemical quenching coefficient of S1401was 0.83,being the smallest.The photosynthetic system of S1401had the weakest self-protection ability.
Dendranthema indicum var.aromaticum;Photosynthetic characteristics;Chlorophyll fluores-cence;Comparative analysis
S682.11
:A
1000-3924(2017)02-013-06
10.15955j.issn1000-3924.2017.02.03
2016-09-19
国家自然科学基金项目(31400590);黑龙江省自然科学基金项目(C2015058)
何淼(1975—),女,博士,副教授,主要从事园林植物种质资源方面的教学科研工作。E-mail:hemiao@nefu.edu.cn
,E-mail:nefuzhouyw@126.com
猜你喜欢
现代盐化工(2021年3期)2021-08-27
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
江苏农业科学(2020年12期)2020-07-20
中国农业科技导报(2020年12期)2020-03-15
食品与生物技术学报(2020年4期)2020-01-06
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
新课程·下旬(2015年10期)2015-10-21
新课程·中旬(2014年10期)2014-12-26
湖北农业科学(2014年13期)2014-08-28
