
shRNA干扰PPARγ基因表达预防兔激素性股骨头坏死的研究*
2017-04-19 03:47冯志尉邓云平郭明刚川北医学院第二临床医学院南充市中心医院南充637000重庆大足区人民医院大足40360
成都医学院学报 2017年2期
冯志尉 ,邓云平,赵 慧,蒲 超,郭明刚,李 伟△.川北医学院第二临床医学院·南充市中心医院(南充 637000);.重庆大足区人民医院(大足 40360)
·论 著·
shRNA干扰PPARγ基因表达预防兔激素性股骨头坏死的研究*
冯志尉1,邓云平2,赵 慧1,蒲 超1,郭明刚1,李 伟1△
1.川北医学院第二临床医学院·南充市中心医院(南充 637000);2.重庆大足区人民医院(大足 402360)
目的 研究激素性股骨头坏死(steroid-induced avascular necrosis of the femoral head,SANFH)的发病机制,观察靶向过氧化物酶体增殖激活受体γ(PPARγ)基因shRNA腺病毒载体预防兔SANFH的作用效果。方法 健康新西兰兔48只,随机分为4组,每组12只。单纯激素组(M组)仅肌注地塞米松;实验组(S组)在肌注地塞米松同时,加用靶向PPARγ基因shRNA腺病毒载体感染兔活体股骨头内骨髓基质干细胞(bone marrow stromal cells,BMSCs);无关序列组(E组)在肌注地塞米松同时,加用搭载无关序列的shRNA重组腺病毒感染兔活体股骨头内BMSCs;对照组(N组)肌注等量生理盐水。测定4组兔血清甘油三酯(TG)、胆固醇(CHO)含量。观察股骨头组织病理学改变。应用RT-PCR 技术检测PPARγ mRNA和Runx2 mRNA表达;应用免疫组化技术检测PPARγ和Runx2蛋白表达情况。结果 M、E、S组血清中TG和CHO含量增高,与N组比较差异均有统计学意义(P<0.05)。M组和E组可见明显骨坏死,脂肪细胞增生肥大,骨小梁变细和稀疏,大量空骨陷凹,S组和N组未见骨坏死及上述改变。S组中PPARγ mRNA和蛋白均呈低表达,Runx2 mRNA及蛋白呈高表达,与M组和E组比较,差异有统计学意义(P<0.05);与N组比较,差异无统计学意义(P>0.05)。结论 靶向PPARγ基因shRNA腺病毒载体可有效阻断BMSCs内PPARγ mRNA表达,抑制其成脂分化;增强Runx2 mRNA表达,促进其成骨分化,预防SANFH的发生,为通过基因靶向治疗SANFH提供新思路。
骨髓基质干细胞;shRNA;过氧化物酶体增殖激活受体γ;激素;股骨头坏死;
激素性股骨头坏死(steroid-induced avascular necrosis of the femoral head,SANFH)为骨科多发病,起病隐匿,致残率高,且缺乏有效防治措施。迄今,国内外研究者针对SANFH的发病机理提出了一系列假说,包括脂质代谢紊乱、血管内凝血、微血管损伤和骨内高压[1-4],但SANFH的确切发病机制尚不清楚。过氧化物酶体增殖激活受体γ(PPARγ)是一种成脂转录因子,在成脂作用和脂肪细胞分化中起着关键性调节作用[5]。研究[6-7]发现,地塞米松能够明显上调鼠、兔、人的骨髓基质干细胞(bone marrow stromal cells,BMSCs)内成脂基因PPARγ的表达,导致BMSCs大量分化为脂肪细胞,造成髓腔内脂肪细胞大量增殖和肥大;髓内压力升高,髓内血窦、毛细血管、静脉受压导致静脉回流受阻,从而影响骨组织血供,最终发生SANFH[8]。本研究采用靶向PPARγ基因的shRNA腺病毒载体感染活体兔股骨头内BMSCs,观察其预防SANFH的作用效果。
1 材料与方法
1.1 材料
1.1.1 实验动物 选取健康新西兰兔48只,雌雄不拘,SPF级,2~3月龄,体质量(2.2±0.5)kg,川北医学院实验动物中心提供。实验过程中对动物处置符合动物伦理学要求。
1.1.2 实验器材及主要试剂 纯化的靶向PPARγ基因的shRNA腺病毒和搭载无关序列的shRNA腺病毒(上海汉恒生物科技公司),PCR引物(上海捷瑞生物公司),地塞米松(山东辰欣药业股份有限公司), Trizol试剂盒(美国OMEGA公司),PCR试剂盒(美国Fermentas公司),RNAKitI提取试剂盒(美国OMEGA bio-tek公司),注射用青霉素钠(美国OMEGA bio-tek公司),血清甘油三酯试剂盒(武汉吉斯生物科技公司),一抗(山羊抗兔PPARγ和Runx2抗体)(北京博奥森生物技术公司),DAB显色试剂盒、辣根过氧化物酶(HRP)标记的二抗和SABC免疫组化试剂盒(武汉博士德生物技术公司),ABI Prism 7300 System (美国Applied Biosystems)倒置相差显微镜(上海万衡精密仪器有限公司)
1.2 方法
1.2.1 实验分组 新西兰兔48只,适应性喂养1周,精确称重后随机分为4组,每组12只。单纯激素组(M组):肌注地塞米松25 mg/(kg·d),连续3 d,作为模型组;实验组(S组):肌注地塞米松25 mg/(kg·d),连续3 d,并于第1天将靶向PPARγ基因shRNA腺病毒载体随机注入兔一侧股骨头内;无关序列组(E组):肌注等量激素,将搭载无关序列的shRNA腺病毒载体随机注入兔一侧股骨头内;对照组(N组):连续3 d肌注0.9%生理盐水25 mg/kg;每次注射时间均在8:00—10:00。
1.2.2 动物模型制作 造模方案[9]:S组兔通过臀肌注射地塞米松,连续3 d,并于实验第1、3周第1天将重组腺病毒注入动物一侧股骨头内。手术方式:1%戊巴比妥钠3 mL/kg麻醉后,随机选择侧别,备皮后常规消毒铺巾,取髋关节后外侧入路,依次切开各层组织,T型切开髋关节囊以暴露股骨头,于头颈交界处以骨穿针与股骨颈呈15°向股骨头内钻入约0.5 cm,用微量注射器取25 μL shRNA腺病毒滴液(109 PFU/μL)通过针道注入股骨头内,用骨蜡封闭穿刺小孔道,逐层缝合。术后连续3 d肌注青霉素40万 U/d预防感染;E组手术方法同S组,注入25 μL搭载无关序列的腺病毒滴液。根据预实验股骨头病理染色结果,各组实验动物分别于术后第4、8周最后1 d分别行空气栓塞分两批处死,每批6只。
1.2.3 血液样本收集及检测 各组动物在实验第4、8周末禁食12 h后,沿耳缘静脉分批次用含肝素锂真空采血管抽取5 mL血液,3 000 r/min离心15 min,分离获得血清样本,使用试剂盒测定各组兔血清中甘油三酯(TG)和胆固醇(CHO)含量。
1.2.4 股骨头组织病理学检测 实验第8周末,手术切取各组兔股骨头标本,首先观察各组股骨头标本的一般特征,再将其沿冠状面一分为二,放入10%的甲醛液中固定24 h,10%EDTA Tris-HCl脱钙7 d,定期更换脱钙液,直至针尖可轻易刺入股骨头,二甲苯透明处理,常规石蜡包埋,制造5 μM厚的冠状面切片,HE染色,应用光镜观察股骨头骨小梁和髓腔结构的变化。使用400倍光学显微镜观察并记录:1)空骨陷凹计数:使用倒置相差显微镜,在HE染色切片中随机选择200个骨陷凹,计算其空骨陷凹所占比率;2)骨坏死发生率=任意一侧股骨头内发生骨坏死的动物数/组内动物总数×100%。参照Matsui等[1]提出的骨坏死程度分级标准进行股骨头组织病理学分级。
1.2.5 股骨头BMSCs中成脂和成骨分化基因表达的检测 将第4、8周收获的股骨头充分研磨匀浆后,用Trizol法提取总RNA,在反转录酶的催化下逆转录合成cDNA,通过荧光定量PCR测定股骨头BMSCs中PPARγ和Runx2的表达情况。RT- PCR 采用 ABI Prism 7300 System ,反应条件为: 94 ℃ 4 min,94 ℃ 20 s,60 ℃ 30 s,72 ℃ 30 s循环35次,72 ℃检测信号,建立PCR产物的扩增曲线。
目的基因引物序列如下:PPARγ 上游5′-AGCAGA
GCAAAGAAGTCGC-3′,下游 5′-CACCTGG TCGTTCAACTCAA-3′;Runx2 上游 5′-AACCC ACGAATGCACTATCCA-3′,下游 5′-GGGACA TGCCTGAGGTGACT-3′;GAPDH 上游 5′- TGC TTCTAGGCGGAGTGTTA-3′,下游 5′- CGTCAC ATGGCATCTCACGA-3′。目的基因相对表达量表示目标基因相对于管家基因(GAPDH)倍数变化的平均值。
1.2.6 免疫组化染色 使用上述已制备的石蜡切片,根据“SABC法”进行免疫组化染色。其中PPARγ和Runx2抗体稀释均为1∶500 ,辣根过氧化物酶(HRP) 标记的二抗(1∶500)37 ℃孵育30 min后, DAB显色。以PBS代替一抗作阴性对照。棕色颗粒代表免疫组化中PPARγ和Runx2的阳性表达,阳性细胞信号定位于细胞膜或细胞质。使用Olympus 光学显微镜观察并拍照。每张切片任意取10个具有代表性的高倍视野(200×)。
1.3 统计学方法
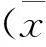
2 结果
2.1 血清学检测结果
实验第4、8周末血清学检查提示:M、E、S组血清中TG、CHO含量增高,与N组比较,差异均有统计学意义(P<0.05)(表1)。


组别第4周第8周TG CHO TG CHO S组3.98±0.68*17.43±2.16**4.97±0.72*21.34±2.02**E组3.95±0.66*17.37±2.01**4.95±0.71*20.78±1.98**M组4.11±0.82**17.61±2.36**5.12±0.91**21.29±2.56**N组1.32±0.192.19±0.321.38±0.222.25±0.37F6.7429.0235.41811.038P0.0010.0000.0040.000
注:与N组相比,*P<0.05;与N组相比,**P<0.01
2.2 股骨头组织病理学观察结果
2.2.1 一般特征 N组股骨头标本外观形态正常,无关节面塌陷及关节囊滑膜增生,关节腔内有少量清亮滑液,关节软骨面平滑且洁净,并显示出健康的颜色(图1A)。激素注射8周后,M、E组股骨头外形正常,无关节面塌陷,可见滑膜轻度增生,关节腔内滑液增多。关节软骨面光滑度较差,但股骨头右上象限表面颜色变深,软骨面可见少许剥脱,软骨下可见散在瘀斑(图1C、图1D)。S组中股骨头外形及表面颜色正常,未见软骨下瘀斑(图1B)。

图1 兔股骨头大体形态
注:黑色箭头:关节软骨面平滑且洁净,并显示出健康的颜色;红色箭头:关节软骨面光滑度较差,股骨头右上象限表面颜色变深,软骨面可见少许剥脱,软骨下可见散在瘀斑
2.2.2 HE染色结果 HE染色后应用倒置相差显微镜观察发现:N组中含有丰富的造血细胞和相对较少的脂肪细胞,形态正常,软骨下骨小梁结构完整,排列规律整齐,骨陷凹中骨细胞清晰可见,细胞核位于中央,且较大,偶见散在空骨陷凹(图2A)。M、E组中可见典型的骨坏死,坏死区域位于临近股骨颈的松质骨和软骨下区,骨髓结构紊乱,骨髓造血细胞变性坏死,坏死的细胞碎屑聚集成团。骨髓脂肪化严重,可见增生肥大的脂肪细胞,部分融合成泡状,骨小梁变细和稀疏,部分断裂成片状,软骨下骨板变薄,骨小梁中含大量空骨陷凹。骨细胞核边聚、固缩,染色差(图2C、图2D)。S组中可观察到少量脂肪细胞,未发现明显的细胞坏死碎屑,骨小梁结构规整,仅见少量空骨陷凹(图2B)。组织学检查表明:N组、M组、E组、S组骨坏死的发生率分别为0.0%(0/6)、100.0%(6/6)、83.3%(5/6)和16.7%(1/6)(表2)。S组与M组和E组间比较,差异有统计学意义(P<0.01),M组和E组间比较,差异无统计学意义(P>0.05),S组和N组间比较,差异无统计学意义(P>0.05)。

图2 第8周各组兔股骨头软骨下区病理组织学变化(HE染色,×200)注:A:N组;B:S组;C:E组;D:M组;蓝色箭头:骨小梁;绿色箭头:空骨陷凹;黑色箭头:脂肪细胞;红色箭头:变性坏死的骨髓造血细胞


组别骨坏死率空骨陷凹率N组 0.04.59±1.46S组16.7*6.54±3.43*E组83.3**62.36±5.16**M组100.0**#67.59±7.24**# 217.40653.927P 0.0010.000
注:与N组相比,*P>0.05,与S组相比,**P<0.01;与E组相比,#P>0.05
2.3 PPARγ和Runx2 mRNA表达结果
第4、8周,组间比较根据Dunnett's t 检验得出,S组中PPARγ mRNA表达量相对于M组和E组降低,差异有统计学意义(P<0.05),S组和N组、M组和E组组间比较,差异均无统计学意义(P>0.05)(表3)。S组中Runx2 mRNA表达量相对于M组和E组升高,差异有统计学意义(P<0.05),S组和N组、M组和E组组间比较,差异无统计学意义(P>0.05)(表3)。


组别第4周第8周PPARγRunx2PPARγRunx2N组1.03±0.070.0057±0.00031.06±0.090.0056±0.0003M组2.46±0.14**#0.0024±0.0005**#2.68±0.21**#0.0021±0.0005**#E组2.39±0.11**0.0028±0.0003**2.57±0.16**0.0026±0.0003**S组1.14±0.11*0.0046±0.0006*1.16±0.13*0.0044±0.0006*F25.4719.95413.0677.286P0.0000.0000.0000.001
注:与N组相比,*P>0.05;与S组相比,**P<0.05;与E组相比,#P>0.05
2.4 免疫组化检测结果
免疫组织化学染色显示:实验第8周,N组股骨头BMSCs中 PPARγ和Runx2表达呈弱阳性(图3A、图4A); M、E组Runx2表达较N组明显减弱(图4C、图4D ),甚至在部分切片中呈阴性表达,PPARγ表达成强阳性; S组中PPARγ和Runx2表达呈弱阳性(图3B、图4B)。该结果证实了上述RT-PCR的分析。

图3 第8周兔股骨头软骨下区免疫组化染色(PPARγ蛋白表达,×200)注:A:N组;B:S组;C:E组;D:M组;N组和S组中股骨头BMSCs中PPARγ蛋白表达呈弱阳性; M、E组PPARγ蛋白表达成强阳性;

图4 第8周兔股骨头软骨下区免疫组化染色(Runx2蛋白表达,×200)
注:A:N组;B:S组;C:E组;D:M组; N组和S组中股骨头BMSCs中Runx2蛋白表达呈弱阳性; M、E组Runx2蛋白表达较N组明显减弱,甚至部分切片表达呈阴性;
3 讨论
激素在临床广泛用于治疗风湿性疾病、自身免疫性疾病和肾脏疾病等,但临床误诊误治、滥用误用激素现象,使SANFH发病率逐年增加,应用糖皮质激素已成为股骨头坏死最常见的危险因素[10]。研究人员对SANFH发病机制提出了多种假设,但目前尚无定论,以致该疾病难以得到有效预防和治疗。目前普遍认同的观点是BMSCs分化失衡导致髓腔内压力增加学说,但所有发病机制均归因于股骨头局部血供的破坏,骨组织的氧分供给减少,导致股骨头局部区域骨小梁和骨髓成分的坏死,最终发生SANFH[11]。
BMSCs可分化为成骨细胞、脂肪细胞、成纤维细胞、软骨细胞及肌细胞等[12]。PPARγ作为具有多个亚型的核转录因子,由相应配体启动后可参与调控BMSCs的增值、分化和凋亡[13]。研究[6-7]发现激素可明显上调鼠、兔、人的BMSCs内成脂基因PPARγ和422aP2mRNA表达,以致BMSCs大量分化成为脂肪细胞。PPARγ被认为是脂肪形成过程中最主要的调控者,当敲除PPARγ基因后成脂分化作用明显减低,且所有亲成脂性细胞信号通路均汇集着PPARγ基因[14]。若上调PPARγ基因表达量,可促使BMSCs向脂肪细胞分化,抑制其高表达可保持BMSCs的成骨分化特性[15-16]。
在SANFH早期阶段,可见脂肪细胞增生和肥大,骨髓造血功能减退,骨细胞内脂质沉积,骨髓坏死,骨小梁变细和稀疏等病理改变。激素可促使髓腔内BMSCs大量向脂肪细胞分化,以致髓腔内压力升高,髓内血窦、毛细血管、静脉受压以致血液回流受到阻碍,从而使骨组织血供受到影响[8];同时伴随大量脂质进入骨细胞内,以致脂质沉积使骨细胞脂肪变性,扰乱骨细胞内脂质代谢的正常进行,导致骨组织脂肪化及变性坏死。
RNAi(RNA interference)是一种高度保守的内源性生物现象[17],其由双链RNA(dsRNA)诱发,通过特异性结合互补链促使同源mRNA高效特异性降解,可被利用成为基因组功能研究的工具[18-19]。应用RNAi技术,可以将基因特异性试剂转染入细胞内,通过序列特异性降解及对mRNA转录产物翻译进行干扰,从而启动基因的敲减,或是基因功能的削弱。RNAi可特异性阻断基因表达,且不改变细胞的生物学行为。该实验构建了针对PPARγ的shRNA腺病毒载体,通过阻断PPARγ基因的表达,抑制股骨头内BMSCs成脂分化,促进其成骨分化,最终起到预防SANFH的作用。本研究给予兔大剂量激素8周后,在M组和E组兔股骨头软骨下区,可见脂肪细胞增生和肥大,骨小梁变细和稀疏,骨髓坏死,空骨陷凹增多等病理学表现。通过RT-PCR和免疫组化技术检测到PPARγ mRNA和蛋白呈低表达,Runx2 mRNA及蛋白呈高表达。但S组未观察到骨坏死的病理学变化,且PPARγ mRNA和蛋白呈低表达,Runx2 mRNA及蛋白呈高表达,兔SANFH发病率降低。
综上所述,靶向PPARγ基因shRNA腺病毒载体可有效阻断BMSCs内PPARγ mRNA表达,抑制其成脂分化,预防SANFH的发生,为通过基因靶向治疗SANFH提供新思路。
[1]Matsui M, Saito S, Ohzono K,etal. Experimental steroid-induced osteonecrosis in adult rabbits with hypersensitivity vasculitis[J]. Clin Orthop Relat Res, 1992(277): 61-72.
[2]Saito S, Inoue A, Ono K. Intramedullary haemorrhage as a possible cause of avascular necrosis of the femoral head. The histology of 16 femoral heads at the silent stage[J]. J Bone Joint Surg Br, 1987, 69(3): 346-351.
[3]Kawai K, Tamaki A, Hirohata K. Steroid-induced accumulation of lipid in the osteocytes of the rabbit femoral head. A histochemical and electron microscopic study[J]. J Bone Joint Surg Am, 1985, 67(5): 755-763.
[4]Wang G, Sweet D, Reger S,etal. Fat-cell changes as a mechanism of avascular necrosis of the femoral head in cortisone-treated rabbits[J]. J Bone Joint Surg Am, 1977, 59(6): 729-735.
[5]Lowell B B. PPARgamma: an essential regulator of adipogenesis and modulator of fat cell function[J]. Cell, 1999, 99(3): 239-242.
[6]王义生. 股骨头坏死的治疗策略与疗效评价[J]. 郑州大学学报(医学版), 2010, 45(4): 533-536.
[7]李子荣,孙伟,屈辉,等.皮质类固醇与骨坏死关系的临床研究[J].中华外科杂志,2005,43(16):1048-1053.
[8]Kawai K, Tamaki A, Hirohata K. Steroid-induced accumulation of lipid in the osteocytes of the rabbit femoral head. A histochemical and electron microscopic study[J]. J Bone Joint Surg Am, 1985, 67(5): 755-763.
[9]Li J, Li Y, Wang Y,etal. Preventive effects of siRNA targeting PPARγ gene on steroid-induced osteonecrosis in rabbits[J]. Connect Tissue Res, 2014, 55(5-6): 322-330.
[10] Liu M, Wang Y S, Li Y B,etal. Construction and identification of the recombinant adenovirus vector carrying a small interfering RNA targeting the peroxisome proliferator-activated receptor-γ[J]. Chin Med J, 2012, 125(4): 671-675.
[11] Childs S G. Osteonecrosis: death of bone cells[J]. Orthop Nurs, 2005, 24(4): 295-301.
[12] Bianco P, Gehron Robey P. Marrow stromal stem cells[J]. J Clin Invest, 2000, 105(12): 1663-1668.
[13] Evans R M, Barish G D, Wang Y X. PPARs and the complex journey to obesity[J]. Nat Med, 2004, 10(4): 355-361.
[14] Tzameli I, Fang H, Ollero M,etal. Regulated production of a peroxisome proliferator-activated receptor-gamma ligand during an early phase of adipocyte differentiation in 3T3-L1 adipocytes[J]. J Biol Chem, 2004, 279(34): 36093-36102.
[15] Li X, Cui Q, Kao C,etal. Lovastatin inhibits adipogenic and stimulates osteogenic differentiation by suppressing PPARgamma2 and increasing Cbfa1/Runx2 expression in bone marrow mesenchymal cell cultures[J]. Bone, 2003, 33(4): 652-659.
[16] Takada I, Kouzmenko A P, Kato S. Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis[J]. Nat Rev Rheumatol, 2009, 5(8): 442-447.
[17] Ketting R F. The many faces of RNAi[J]. Dev Cell, 2011, 20(2): 148-161.
[18] Mohr S, Bakal C, Perrimon N. Genomic screening with RNAi: results and challenges[J]. Annu Rev Biochem, 2010, 79: 37-64.
[19] Perrimon N, Ni J Q, Perkins L. In vivo RNAi: today and tomorrow[J]. Cold Spring Harb Perspect Biol, 2010, 2(8): a003640.
A Study on the Preventive Effect of the Interference of PPARγ Gene Expression by shRNA on Steroid-induced Avascular Femoral Head Necrosis in Rabbit Models
FengZhiwei1,DengYunping2,ZhaoHui1,PuChao1,GuoMinggang1,LiWei1△.
1.SecondClinicalSchoolofNorthSichuanMedicalCollege&NanchongCentralHospital,Nanchong637000,China; 2.DazuDistrictPeople’sHospitalofChongqing,Dazu402360,Chongqing
Objective To investigate the pathogenesis of steroid-induced avascular necrosis of the femoral head (SANFH), and explore the preventive effect of shRNA targeting PPARγ gene on SANFH in rabbit models. Methods Forty-eight healthy New Zealand rabbits were randomly into the four groups including Group M treated with dexamethasone only, Group S with dexamethasone and a recombinant adenovirus vector carrying shRNA targeting PPARγ gene and infecting the rabbits’ bone marrow stromal cells (BMSCs), Group E with dexamethasone and a vector carrying irrelative sequence of shRNA adenovirus vector infecting BMSCs, and Group N with Saline as the control group. The levels of serum triglycerides (TG) and cholesterol (CHO) in the four groups were detected and the pathological changes of femoral head were observed. RT-PCR was used to assay the expression levels of PPARγ mRNA and Runx2 mRNA, and the immunohistochemistry was performed to detect the expression of PPARγ and Runx2. Results The contents of serum TG and CHO in Groups M, E and S increased, and they were significantly different from those in Group N (P<0.05). Both Group M and Group E showed obvious osteonecrosis and changes such as adipocyte proliferation and hypertrophy, thinner and sparse trabeculae, and plenty of empty osteocyte lacunae, while the osteonecrosis and changes were not found in Group S and Group N. The expression levels of PPARγ mRNA and protein were low while the expression levels of Runx2 mRNA and protein were high in Group S. The expression levels of Group S were significantly different from those of Group M and Group E respectively (P<0.05), while they were not significantly different from those of Group N (P>0.05). Conclusion The adenovirus vectors shRNA targeting PPARγ gene can effectively inhibit the expression of PPARγ mRNA in BMSCs and the adipogenic differentiation, promote osteogenic differentiation of BMSCs, and finally prevent steroid-induced osteonecrosis in rabbits, which may provide a novel approach to the treatment of steroid-induced osteonecrosis by means of gene targeting therapy.
Bone marrow stromal cells; ShRNA; Peroxisome proliferator-activated receptor gamma; Glucocorticoids; Osteonecrosis of femoral head
*通信作者:四川省教育厅自然科学基金(No:14ZA0185)
http://kns.cnki.net/kcms/detail/51.1705.R.20170327.1001.008.html
10.3969/j.issn.1674-2257.2017.02.003
R681.8
A
△通信作者:李伟,E-mail: subjudice@126.com
猜你喜欢
传染病信息(2022年3期)2022-07-15
音乐教育与创作(2020年9期)2021-01-04
浙江医学(2019年9期)2019-05-20
灾害医学与救援(电子版)(2018年1期)2018-06-05
今古传奇·故事版(2017年23期)2018-01-12
上海农业学报(2017年4期)2017-04-10
现代检验医学杂志(2016年1期)2016-11-12
中华骨与关节外科杂志(2016年6期)2016-05-17
中国卫生标准管理(2015年10期)2016-01-15
故事会(2016年1期)2016-01-05
