
宿根蔗苗期大螟幼虫空间动态的地统计学分析
2017-04-19 03:23KH3D罗志明尹炯黄应昆单红丽李文凤王晓燕张荣跃
西南农业学报 2017年1期
[KH-3D]罗志明,尹炯,黄应昆,单红丽,李文凤,王晓燕,张荣跃
(云南省农业科学院甘蔗研究所,云南开远661699)
宿根蔗苗期大螟幼虫空间动态的地统计学分析
[KH-*3D]罗志明,尹炯,黄应昆*,单红丽,李文凤,王晓燕,张荣跃
(云南省农业科学院甘蔗研究所,云南开远661699)
大螟是危害甘蔗较为严重的一类钻蛀性害虫。应用地统计学分析方法,研究了宿根蔗苗期大螟幼虫田间分布规律与扩散动态,建立了不同调查时间在0°、45°、90°、135°等4个方向上大螟幼虫的半变异函数及理论半变异函数,并用Kriging插值法对空间分布数据进行插值和模拟。结果表明,大螟幼虫密度呈单峰型增长,聚集度和随机变异随虫口密度的增加而减小;大螟幼虫呈聚集型分布,随着虫口密度的升高,聚集度降低,相关性增强,范围在5.33~43.59 m;Kriging插值表明蔗苗前期各斑块间相关性弱,蔗苗中后期各斑块间相关性强。
宿根蔗;大螟;地统计学;空间动态
大螟Sesamia inferens Walker,属鳞翅目,夜蛾科,是危害甘蔗较为严重的一类钻蛀性害虫[1]。大螟1年发生5~6代,由于其具有危害期长、繁殖快、钻蛀危害等特点,必须注重早期的预警监测及第一、二代的防治工作[2]。空间分布是昆虫种群的重要特征之一,是昆虫种群的生物学特性与特定环境条件相互作用的结果[3],研究甘蔗苗期大螟幼虫的空间分布有助于了解大螟在甘蔗苗期的生态特性,对研究大螟种群消长规律、预测未来的发展趋势,指导科学有效防控大螟具有重要意义。地统计学(Geostatistics)是在地质分析和统计分析互相结合的基础上形成的一套分析空间相关变量的理论和方法,主要研究在空间分布上既具有随机性,又具有结构性的自然现象,它能最大限度地利用空间数据,揭示各种自然现象的本质,比经典统计学方法更精确,可有效避免系统误差[4-5]。
采用经典生物统计学方法研究大螟在不同作物上的分布扩散规律已有报道。尹炯等[6]利用经典统计学方法研究了大螟枯心苗的空间分布型及抽样技术;苏艺等[7]分析了海南橡胶园砖红壤光谱反射特征;罗志明等[8]研究了大螟和黄螟在蔗苗上的生态位及其种间竞争关系;韩兰芝等[9]研究了大螟幼虫田间扩散及成虫飞行能力;敬甫松[10]研究了大螟在玉米田的空间格局及其抽样技术;覃永嫒等[11]研究了鲜食玉米桂糯518的施肥情况。地统计学在昆虫种群结构研究中已有广泛的应用,Liebhold等[12]根据不同尺度的变差图,证明了舞毒蛾卵块在0.025~50 km内聚集;Kemp等[13-14]研究证明了蝗虫在1~100 km的距离存在空间相关性;闫香慧等[15]研究了白背飞虱和褐飞虱种群的空间分布特点;石根生等[16]研究了马尾松毛虫空间格局;巫厚长等[17]研究了烟蚜及其捕食性天敌草间小黑蛛种群空间结构;夏鹏亮等[18-19]研究了恩施烟区烟夜蛾和桃蚜在烤烟田的空间动态,明确了其在各烤烟生育区的空间格局;黄寿山等[20]应用地统计学分析了二化螟越冬幼虫空间分布图式。
甘蔗种植制度多样,总体可分为新植蔗和宿根蔗,从种植面积来看,宿根蔗占整个种植面积的60 %以上[21]。大量的田间调查发现,新植蔗大螟虫源主要来自于周边蔗园(少量蔗种自带),而宿根蔗则是蔗园内上年残留虫口。昆虫种群空间动态受虫源影响[22],不同的虫源决定着不同的空间分布和危害水平。本研究应用地统计学方法对宿根蔗园大螟幼虫空间格局进行分析,以期探明其空间聚集程度和时间扩散动态,为预测报和综合防治提供理论依据。
1 材料与方法
1.1 调查方法
调查设于云南省农业科学院甘蔗研究所试验示范基地(N 23°42'39.9″,E 103°15'43.7″),海拔1 050 m,属水田蔗区。1年宿根,甘蔗品种为云蔗05-211,种植行距1 m。将864 m2(18 m×48 m)的调查田按1 m×2 m格式分成432个2 m2的样方(点),每样方全部调查,以大螟危害新增枯心苗数作为样点虫口密度指标,并按样方位置分别记录。调查自2012年3月28日开始,直到5月28日甘蔗大培土前结束。每7~8 d 1次,共调查9次。调查日期依次为3月28日、4月5日、4月12日、4月20日、4月28日、5月6日、5月13日、5月21日和5月28日。
1.2 数据分析方法
基础数据用EXCEL统计,半变异函数计算、理论半变异函数模型拟合和Kriging插值模拟用DPS 14.50软件完成。
应用地统计学中最常用的半变异函数来反映大螟幼虫空间格局及空间相互依赖关系,同时对得到的变异函数用不同的理论模型进行拟合,通过拟合模型中最重要的3个参数块金常数、变程和基台值来反映大螟幼虫种群分布随机部分的空间异质性和在某一范围内存在空间相关性等。根据调查数据,以东-西为0°方向,分别按逆时针旋转45°、90°、135°,共设置4个方向进行理论半变异函数拟合,确定最佳理论拟合模型,并计算出相应的块金值、基台值和变程等参数[23-24]。
空间分布的区域化变量用Kriging插值法生成分布图,它是基于区域化变量理论,当获得了某个变量的半变异函数的模拟模型后,可利用样点观测值对研究区域上未取样点的区域化变量值进行最小误差估计[23]。
2 结果与分析
2.1 大螟幼虫空间分布数据的统计特征
不同时间大螟幼虫数据的统计特征见表1。3月底至5月底甘蔗苗期,大螟幼虫密度呈单峰型增长,峰值出现在5月6日和5月13日,平均密度均为2.24头/点,最大值14头/点。大螟幼虫的平均密度和标准差4月12日前较小,4月28日至5月28日期间,变异系数前者较大,后者较小。从平均值与最大值的变化趋势来看,两者基本一致,但两者增幅不同,平均值增幅大于最大值增幅,表明其空间分布的不均匀性随密度的增大而减小,种群密度的增大一定程度上提高了种群结构的稳定性。从峰度值和偏度值来看,各调查时间大螟幼虫数据均为右偏的分布(偏度>0),偏度越大相应的峰度越高,且随调查时间的后移,偏度逐渐减小,表明随着调查时间后移,大螟幼虫空间分布聚集度降低。
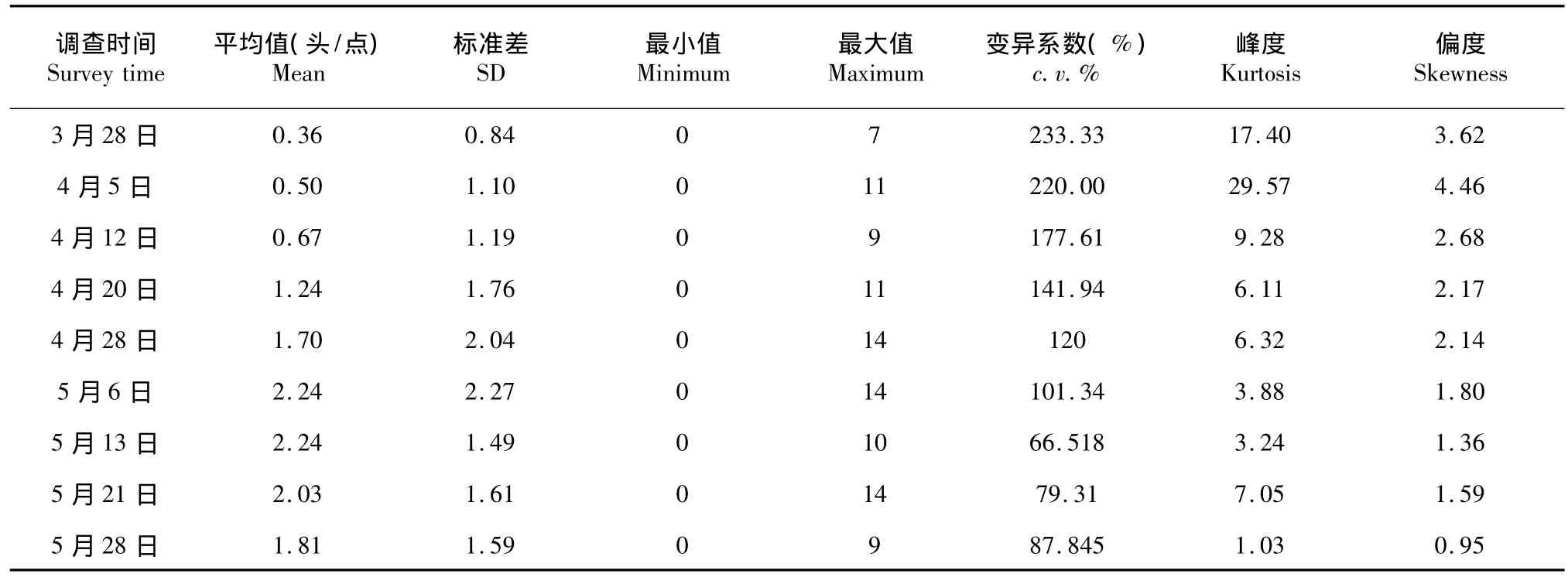
表1 大螟幼虫空间分布的描述性统计Table 1The descriptive analysis of spatial distribution of S.inferens larvae

表2 不同时期大螟幼虫在不同方向上空间特点及模型参数Table 2Model parameters and spatial patterns of S.inferens larvae in different direstion at the different seedling stages of ratoon cane

各图横坐标为东向西方向,纵坐标为北向南方向,数值表示样方位点Abscissa meant the direction of east to west;Ordinate meant the direction of north tosouth;Numerical meant quadrat site图1大螟幼虫种群Kriging插值分布Fig.1Kriging of spatial pattern of S.inferens larvae
2.2 大螟幼虫密度的空间变异
根据2012年3-5月的9次调查数据,分别计算不同时间大螟幼虫密度在0°、45°、90°、135°方向的实际半变异函数,然后利用球形模型、指数模型、高斯模型、球形+球形套合模型、球形+指数模型和线性有基台模型来拟合4个方向上的半变异函数理论模型,确定最佳理论拟合模型,并计算出相应的块金值、基台值和变程等参数,结果见表2。
从表2可看出,4月12日前,在4个方向上半变异理论拟合模型均为球形模型,决定系数除少数在0.7~0.8,其余均超过0.8,拟合度较好,属聚集型分布;4月20日至5月28日,各方向上理论模型以球形模型和指数模型为多,并伴有线性有基台模型和随机分布,且大多拟合决定系数低于0.7,拟合效果差,分布型复杂。
块金值、基台值和变程等3个参数,不同时间不同方向的参数值有着各自的特性,从4个方向的平均值来看,块金值(9个时间的平均值分别为0.76、1.58、4.26、7.76、3.08、2.20、1.21、2.39、2.19)、基台值(9个时间的平均值分别为0.38、1.30、1.21、2.78、10.84、9.90、2.28、2.73、2.58)和变程(9个时间的平均值分别为5.33、7.90、8.59、17.54、43.59、33.72、13.42、16.51、16.68)均与田间虫口密度变化趋势相似,虫口密度低时数值低,随着虫口密度的增加数值逐渐加大。5月13日后,虫口密度稳定在1.81~2.24头/点,基台值稳定在2.28~2.73,变程稳定在13.42~16.68 m。
在随机程度上,4月12日前,随机程度均大于0.75,表明此时空间相关性较弱;4月20-28日,各方向随机程度差异较大,此时空间相关性随不同方向变化较大;5月6-13日,随机程度低于0.75,部分方向还低于0.25,表现出较强的空间相关性;5月21-28日,随机程度大于0.75,空间分布随机性增加。
2.3 大螟幼虫种群的空间分布模拟
由图1可看出,随着调查时间后移,大螟种群聚集斑块数量增多,并以长条形斑块状向周围扩散。其中,3月28日至4月12日,斑块数量虽有增加,但斑块间的重合度较差,表明此期间新增斑块与以前斑块间相关性较差;4月20日以后斑块数量和面积逐渐增加,且相互间的斑块重合度高,表明此期间大螟种群空间分布相关性强,且不同调查时间的斑块间相关性也较强。从种群密度的变化趋势来看,随着种群密度的增加,大螟种群聚集斑块数增加,当种群密度达最大值时(5月6日、5月12日)聚集斑块已遍布蔗园各处,随后,斑块面积逐渐扩大,种群分布趋于均匀。
3 讨论
蔗苗受大螟危害后形成枯心苗,以大螟危害枯心苗代替虫口数量具有一定的代表性[25]。研究发现,蔗苗前期大螟虫口密度低,随着蔗苗的生长,田间虫源得到了积累,再加上大螟世代数的增加,繁殖系数不断提高,虫口数量随之增加,体现了一般昆虫发生的普遍规律。虫口密度的最大值与平均值的变化趋势完全一致,且虫口密度的变化幅度大于最大值变化幅度,表明同一田块大螟幼虫空间分布随时间变化均匀性提高。大螟幼虫发生动态呈单峰型,峰值出现在5月6日和5月13日2个连续调查日,可能与这2个时段间正值大螟第2代幼虫盛发期[1]有关。从变异系数、峰度和偏度来看,随着调查的后移,数值逐渐减少,表明蔗苗前期大螟幼虫聚集度高,随着蔗苗的生长,大螟聚集度降低,种群结构趋于稳定。
应用地统计学方法进行半变异模型拟合,4月12日将整个甘蔗苗期大致划分为2个阶段,即4月12日前,在4个方向上半变异理论拟合模型均为球形模型,且决定系数大,表现出较强的聚集性;4月12日后,各方向上理论模型以球形模型和指数模型为多,并伴有线性有基台模型和随机分布,且大多拟合决定系数低,拟合效果差,分布型复杂,这与大螟年度发生规律有关。4月中旬为大螟1代幼虫发生高峰期,在此之前由于发生虫源单一,分布型较一致,但随着时间的后移,1、2代大螟重叠发生,加上外来虫源的进入,田间分布趋于复杂。在4个方向的平均基台值和平均变程等参数方面,它们均随虫口密度的增加而增加,直到蔗苗后期稳定在一定范围,即基台值稳定在2.28~2.73,变程稳定在13.42~16.68 m,这反映了大螟区域化变量的一个特征。
大螟幼虫田间分布主要受虫源影响。通过Kriging插值获得大螟幼虫不同时间的田间分布模拟图,从图上可以清晰反映出各自空间分布特征。在蔗苗前期(3月28日至4月12日)斑块数量较少,所包含面积小,且各调查点间重合度较差,在蔗苗中后期(4月20日以后)斑块数量多,所包含面积大,且相互间的斑块重合度较高,进一步说明了蔗苗前期大螟幼虫的聚集性和不稳定性。
[1]黄应昆,李文凤.现代甘蔗病虫草害原色图谱[M].北京:中国农业出版社,2011.
[2]罗志明,黄应昆,李文凤,等.高原生态蔗区甘蔗螟虫猖獗原因与防治对策[J].动物学研究,2009(昆虫学专辑):105-109.
[3]周国法,徐汝梅.生物地理统计学[M].北京:科学技术出版社,1998.
[4]Rossi R E,Mulla D J,Journel A G,et al.Geostatistical tools for modeling and interpreting ecological spatial dependence[J].Ecological Monographs,1992,62(2):277-314.
[5]周强,张润杰,古德祥.地质统计学在昆虫种群空间结构研究中的应用概述[J].动物学研究,1998,19(6):482-488.
[6]尹炯,黄应昆,李文凤,等.大螟蔗田枯心苗的空间分布型及抽样技术研究[J].植物保护,2012,38(4):127-130.
[7]苏艺,杨磊,林清火,等.海南橡胶园砖红壤光谱反射特征分析[J].南方农业学报,2015,46(3):385-390.
[8]罗志明,申科,黄应昆,等.大螟和黄螟在蔗苗上的生态位及其种间竞争[J].应用昆虫学报,2014,51(4):1046-1051.
[9]韩兰芝,彭于发,吴孔明.大螟幼虫田间扩散及成虫飞行能力研究[J].植物保护,2012,38(4):9-13.
[10]敬甫松.大螟在玉米田的空间格局及其抽样研究[J].昆虫知识,1987,24(2):71-78.
[11]覃永嫒,时成俏,王兵伟,等.鲜食玉米桂糯518的肥料效应试验及推荐施肥分析[J].南方农业学报,2014,45(2):244-249.
[12]Liebhold A M,ZhangX,Hohn M E.Geostatistical analysis of gypsymoth(Lepidoptera:Lymantriidae)egg mass populations[J].Environmentul Entomology,1991,20(5):1407-1417.
[13]Kemp W P.Probabiling of outbreak for Rangeland grasshoppers (Orthoptera:Acrididae)in Montana:application of Markwvian principle[J].Environmental Entomology,1987,80(6):1100-1105.
[14]Kemp W P,Kalaris T M,Quimby W F.Rangelandgrasshopper (Orthoptera:Acrididae)spatial variability:macroscalepopulation assessment[J].Environmental Entomology,1989,82(5):1270-1276.
[15]闫香慧,赵志模,刘怀,等.白背飞虱若虫空间格局的地统计学分析[J].中国农业科学,2010,43(3):497-506.
[16]石根生,李典谟.马尾松毛虫空间格局的地学统计学分析[J].应用生态学报,1997,8(6):612-616.
[17]巫厚长,徐光曙,房明惠,等.烟蚜及其捕食性天敌草间小黑蛛种群空间结构分析[J].应用生态学报,2004,15(6):1039-1042.
[18]夏鹏亮,王瑞,谭军.湖北恩施烟夜蛾幼虫田间分布格局的地统计学分析[J].应用生态学报,2014,25(3):825-830.
[19]夏鹏亮,王瑞,王昌军,等.恩施烟区无翅桃蚜在烤烟田空间动态的地统计学分析[J].生态学报,2014,34(5):1198-1204.
[20]黄寿山,胡慧建,梁广文.二化螟越冬幼虫空间分布图式的地理统计学分析[J].生态学报,1999,19(2):250-253.
[21]江永,陈凯,黄毓芬.宿根甘蔗生长特性和产量结构特点[J].中国糖料,2004(2):22-25.
[22]张孝羲.昆虫生态及预测预报[M].北京:中国农业出版社,1997:53-69.
[23]唐启义.DPS数据处理系统[M].北京:科学出版社,2013: 1113-1132.
[24]王政权.地质统计学在生态学中的应用[M].北京:科学出版社,1999.
[25]潘杰,王涛,宗世祥,等.红脂大小蠹种群空间格局地统计学分析及抽样技术[J].生态学报,2011,31(1):195-202.
(责任编辑 王家银)
Geostatistical Study on Spatial Dynamics of Sesamia inferens Walker Larvae in Ratoon Cane Seedling Stage
LUO Zhi-ming,YIN Jiong,HUANG Ying-kun*,SHAN Hong-li,LI Wen-feng,WANG Xiao-yan,ZHANG Rong-yue
(Sugarcane Research Institute,Yunnan Academy of Agricultural Sciences,Yunnan Kaiyuan 661699,China)
Sesamia inferens Walker was a borer pests seriously damaging sugarcane.The spatial pattern and dispersal dynamic of S.inferens Walker larvae were studied in ratoon cane at the seedling stage,and the semivariograms and theoretical semivariogams of S.inferens larvae in the four directions(0°,45°,90°and 135°)were established based on the geostatistical method by using the survey data in the different periods.Isoclines maps of the S.inferens larvae at each stage were plotted by using Kriging interpolation.The results showed that the density of S.inferens larvae showed a unimodal increase,the aggregation and the random variation decreased with the increasing of the density.The population of S.inferens larvae showed an overall clumped distribution,the aggregation decreased and the correction increased with a range of 5.33-43.59 m with the density increased.Kriging interpolation indicated that the corrections between the patches were at the early seedling stage and were strong at the intermediate and late seedling stage.
Ratoon cane;Sesamia inferens Walker;Geostatistics;Spatial dynamics
Q968.1
A
1001-4829(2017)1-0105-06
10.16213/j.cnki.scjas.2017.1.019
2015-03-17
现代农业产业技术体系建设专项资金(CARS-20-2-2);云南省现代农业产业技术体系建设专项资金;云南省科技计划项目(2013BB013);云南省农业科学院院基金(2014CZJC010)
罗志明(1973-),男,云南华宁人,助理研究员,主要从事甘蔗病虫害防治研究,kylzm@163.com,*为通讯作者,huangyk64@163.com。
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2021年3期)2021-12-05
今日农业(2021年9期)2021-07-28
今日农业(2021年4期)2021-06-09
天津农林科技(2021年2期)2021-04-16
湖南电力(2021年1期)2021-04-13
信阳农林学院学报(2021年1期)2021-04-01
林业科技(2020年3期)2021-01-21
红土地(2018年7期)2018-09-26
农村农业农民·B版(2016年7期)2016-10-21
