
温度和盐度对大竹蛏孵化及幼虫生长变态的影响
2016-08-30 05:49郝咏芳曹琛邢佐平黄金勇匡少华宋松伟
水产养殖 2016年8期
郝咏芳,曹琛,邢佐平,黄金勇,匡少华,宋松伟
(1.营口市水产科学研究所,辽宁 营口 115004;2.大连海洋大学,辽宁 大连 116023)
温度和盐度对大竹蛏孵化及幼虫生长变态的影响
郝咏芳1,2,曹琛1,邢佐平1,黄金勇1,匡少华1,宋松伟1
(1.营口市水产科学研究所,辽宁 营口 115004;2.大连海洋大学,辽宁 大连116023)
大竹蛏(Solen grandis Dunker)隶属瓣鳃纲,异齿亚纲,帘蛤目,竹蛏科,竹蛏属。贝壳较长,呈竹筒状,一般壳长约为壳高的4~5倍,足极发达。营埋栖生活,生活在潮下带至水下20 m水深处,生存底质为沙底、泥沙底,主食浮游性较弱易于下沉或底栖的硅藻类[1]。大竹蛏广泛分布于中国、朝鲜半岛、日本以及菲律宾沿海,其肉质营养价值高,蛋白质含量高,必需氨基酸种类齐全,呈味氨基酸比例高,脂肪中含有大量的EPA及DHA等不饱和脂肪酸[2-3]。
目前,有关大竹蛏的研究主要集中在繁殖生物学[4-6]、人工育苗[7-10]等方面,培育密度及饵料种类[11]、环境因子[12-13]、盐度等对大竹蛏稚贝生长发育影响的研究[14]也有报道,但温度及盐度对大竹蛏卵的孵化及幼虫变态影响的相关研究尚未发现。笔者进行这方面的试验,为大竹蛏人工育苗幼体孵化及变态阶段积累基础数据,以期为其苗种的规模化培育技术示范及推广提供理论依据。
1 材料与方法
1.1材料
1.1.1试验场地及主要设施场地位于营口现代渔业科技产业园贝类研发车间。天然海水沉淀、砂滤后使用,盐度22,pH值7.5~8.6,水温17~18℃,水质符合我国渔业水质标准[3]。试验用容器为35 cm×50 cm×15 cm的白塑料槽。
1.1.2试验用大竹蛏受精卵及D型幼体大竹蛏种贝购自营口市白沙湾海区,为2~3龄成熟大竹蛏亲贝,个体健壮、生长旺盛、无损伤、肥满度高、壳长8 cm以上。种贝暂养至性腺成熟自然产卵,300目筛绢网收集同一批受精卵及250目筛绢网选育出的同一批D型幼体。
1.2方法
1.2.1温度对幼虫孵化及生长变态的影响试验海水盐度22,水温在15~36℃范围内,3℃为一个梯度,设置8个组。以种蛏产卵时的水温为基点,按每10 min升高或降低2℃的速率通过冰袋降温或加热棒升温调节,控温仪控制,温差不超过±0.1℃。1.2.2盐度对幼虫孵化及生长变态的影响试验海水水温23~24℃,盐度6~38,以4为梯度,共设9个盐度组。试验用水由盐度100的浓缩海水、盐度22的天然海水、充分曝气的自来水按比例配制而成,用YSI水质快速分析仪标定。不同试验组海水一次配制,分装于20 L纯净水塑料桶中保鲜膜封口备用。以突变方式调节各组盐度,盐度对变态率影响的试验时间为28 d。
1.2.3日常管理2014—2015年生产和试验同步进行。每个试验组设置两个平行,各组微量充气,保证溶氧,自然光照1 000 lx以内。孵化试验受精卵密度4个/mL,变态试验D型幼体密度2个/mL。试验在装有2 L不同梯度水温、盐度海水的塑料槽中进行。定时取样观察、镜检、记录幼虫的发育情况及各阶段形态变化和幼虫数量。
孵化率试验时间较短,要定时观察记录。变态率试验中,日换水1次,换水量70%;日投饵2次,初期金藻(Isochrysis zhanjiangensis)开口,后与角毛藻(Chaetoceros sp)搭配投喂,前期投喂密度2 000 cell/mL,随着各组幼体生长发育和数量的变化情况适当调整投喂量,以保持水体单胞藻细胞密度2 000 cell/mL以上,从而保证其足量摄食。镜检幼虫存活、变态情况,测量壳长,及时捞出死亡幼体(死亡的界定以贝壳张开为标准)。试验结束时,统计各组幼体孵化率、变态率和壳长日增长率。
1.3数据统计及处理
孵化试验:受精卵孵化为D形幼虫所需时间为孵化时间,凡D形幼虫出现壳变形扭曲、左右壳不对称、铰合线弯曲、壳缘有缺刻、面盘或纤毛异常者均计为畸形个体,不计入孵化率。
变态试验:大竹蛏的个别D型幼体壳长230 μm时即开始变态,发育至眼点圆大、面盘完全脱落、足形成可爬行时视为完成变态,此时幼体壳长大多在250~300 μm之间,此阶段所需时间为变态时间,面盘及外壳异常者记为畸形个体,不算入变态率。计算公式如下:
孵化率:Rh=dh1/dh0×100%
变态率:Rs=ds1/ds0×100%
壳长平均日增长量:Qg=(L1-L0)/(t1-t0)
式中:dh1为D型幼体数量,dh0为受精卵数量;ds1和ds0分别表示截止试验结束成功变态幼体密度和初始D型幼虫数量;L1和L0分别为试验结束时幼体壳长和初始D型幼虫壳长(μm);t1和t0分别为试验结束时间和D型幼体形成时间。试验期间的死亡个体仅用于计算病死率指标。
试验数据采用平均值(M)±标准差(SD)表示;Excel(2003)对数据进行处理、作图,SPSS17.0对数据进行单因素方差分析(ANOVA),用于检验处理间的差异显著性(P<0.05)。
2 结果与分析
2.1温度对大竹蛏幼虫孵化及生长变态的影响
温度对大竹蛏幼虫孵化及生长变态的影响见图1、图2。
试验结果表明,水温对于大竹蛏受精卵孵化率影响具有统计学意义(P<0.05)。大竹蛏受精卵对低温比对高温适应性略强。水温12℃,幼体分裂至多细胞即停止发育或畸形,无法完成孵化;15℃时,受精卵胚胎发育进程缓慢,大部分发育畸形,只有28%的受精卵可以正常孵化,且所需时间较长;15~24℃时,随着水温升高,受精卵孵化至D型幼虫时间缩短,孵化率显著升高(P<0.05);水温24℃时,孵化率最高为79.25%,略高于27℃的72.75%,变态所需时间也最短,仅为5 d;27℃起,水温上升孵化时间逐步增长,孵化率降低显著(P<0.05);33℃时受精卵大部分发育至多细胞便开始解体,少数畸形,孵化率仅15.25%,但孵化时间和18℃相同,均略低于15℃;36℃时极个别幼虫能发育至担轮幼虫,但也无法正常孵化。因此,大竹蛏受精卵孵化的适宜温度范围18~30℃,最适温度范围21~27℃,在此范围内,幼虫孵化较快且孵化率较高。因此,大竹蛏受精卵孵化过程中,应将水温控制在21~27℃。
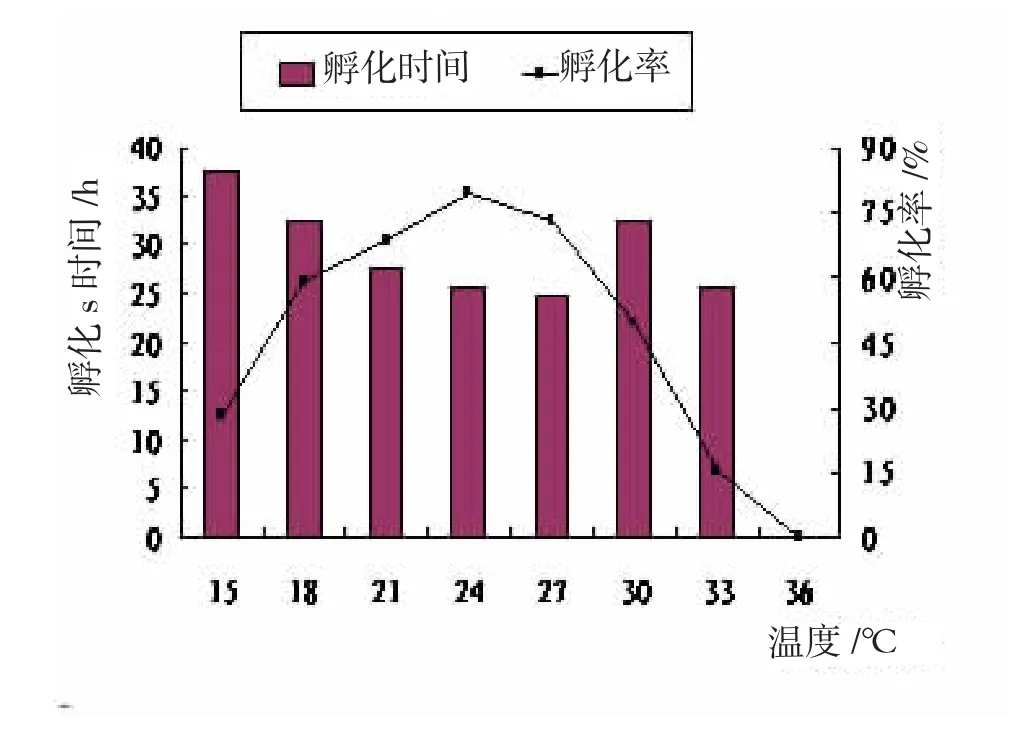
图1 温度对幼体孵化的影响

图2 温度对幼体变态的影响
水温对大竹蛏幼体变态率的影响显著(P<0.05),12℃无法完成变态,少数发育至壳顶期即死亡;15℃时D型幼体活力弱,摄食差,部分幼体面盘伸缩异常,48 h开始出现面盘解体,72 h后有死亡个体,变态时间较长,变态率仅39.75%;15~24℃时,随着温度上升,D型幼体壳长日增长增大极显著,变态率显著上升,24℃时壳长日增长23.4 μm/ d,变态率85.45%;27℃壳长日增长、变态时间及变态率与24℃差异均无统计学意义(P>0.05);水温30℃时,12 h内幼虫表现异常活泼,面盘伸缩频繁,滤食能力强,但此后逐渐出现沉底、纤毛脱落等现象,壳长日增长量下降显著(P<0.05);36℃,12 h后幼虫空胃率70%以上,几乎停止发育,面盘解体、逐渐沉底,无法完成变态。因此,大竹蛏D型幼体变态的适宜温度范围18~30℃,最适温度范围21~27℃。
2.2盐度对大竹蛏幼虫孵化及生长变态的影响
如图3所示,水温23~24℃时,盐度对于大竹蛏受精卵的孵化率及发育速度均影响显著(P<0.05)。盐度过高或过低,受精卵无法正常孵化,盐度为6时大竹蛏受精卵分裂至原肠期终止,盐度38时受精卵也无法完成孵化;盐度10时,大部分幼虫发育至担轮幼体即终止,孵化率仅2.75%,且孵化时间较长;盐度10~34,各组均有D型幼虫孵出,但孵化率差异具有统计学意义(P<0.05);盐度10~26时,随着盐度升高孵化率显著升高(P<0.05),孵化时间显著降低(P<0.05);盐度26孵化率最大为82.25%,且孵化时间最短,盐度22次之;盐度26~34,盐度升高孵化率显著下降(P<0.05),孵化时间显著增长(P<0.05);盐度34时受精卵多数发育至多细胞,分裂球分裂畸形,最终死亡,孵化率仅13.85%。由此可见,大竹蛏受精卵孵化适宜盐度范围为18~30,最适范围为22~30,在此范围内,大竹蛏的孵化率较高,发育速度较快,初孵D形幼虫活力较好。
由图4可知,盐度变化对大竹蛏幼体的变态率影响显著(P<0.05)。盐度过高、过低,大竹蛏幼虫无法正常变态。盐度6时,D型幼体发育12 h后陆续闭壳沉底,镜检发现幼虫面盘纤毛和胃部仍在活动,60 h后全部死亡;盐度10时变态所需时间较长,变态率仅3.5%;盐度14~26,随着盐度升高幼体变态率显著增大(P<0.05),摄食、活力较强,生长速度也逐渐提高;盐度26时壳长平均日增长量和变态率最大,变态时间最少,变态规格相对较小(壳长231 μm),盐度22时次之;盐度26~34,变态率随盐度上升显著下降(P<0.05),生长发育减慢,变态时间增长;盐度38时,幼虫高渗反应明显,游动异常,24 h出现死亡,3 d后病死率40%,存活的幼虫以壳顶中期为主,6 d后全部死亡;各盐度组对大竹蛏幼体变态率的影响依次为:盐度26>盐度22>盐度30>盐度18>盐度34>盐度14>盐度10。因此,大竹蛏D型幼体变态的适宜盐度范围为18~30,最适盐度范围22~30。

图3 盐度对幼体孵化的影响

图4 盐度对幼体变态的影响
3 讨论
3.1温度大竹蛏幼虫孵化及变态的影响
贝类属于变温动物,其体温及代谢随环境温度的改变而变化。温度作为影响贝类生长发育最重要的环境因子之一,在一定范围内缓慢提高可以促进贝类的生长发育。本研究表明,大竹蛏受精卵孵化和变态的适宜温度为18~30℃。低温时胚胎发育缓慢且畸形率高,无法正常孵化或顺利完成变态。最适温度为21~27℃,在此范围内,胚胎发育正常且发育较快,孵化率和变态率较高。这与陈爱华[12]的大竹蛏稚贝存活和生长的适宜温度范围6~30℃,最适温度范围21~30℃略有不同,可能是由于大竹蛏受精卵和浮游幼虫较稚贝胚胎发育不完善,对低温抵抗力差有关。与青蛤胚胎、浮游幼虫的适宜温度24~32℃,最适生长温度26~30℃[15]不同,与毛蚶孵化的适宜温度25~30℃[16]、幼虫生长最适温度30℃也不同,这可能与它们的生活习性有关,大竹蛏在辽东湾一带5月下旬至6月为繁殖盛期,此时天然海水温度范围在18~22℃;而青蛤繁殖盛期是每年的7月至9月,正处于盛夏季节[15],海水温度较高;毛蚶的产卵大多在6月底至7月份,海水温度也较高。因此大竹蛏幼体的最适孵化及变态温度范围应该在21~27℃。
3.2盐度对大竹蛏幼虫孵化及变态的影响
海洋贝类属于变渗动物,能够根据周围环境海水盐度不同而调节其渗透压,以利于摄食和生长[12]。贝类对盐度的耐受力与渗透调节能力、营养状况和遗传变异(包括抗逆基因的表达)有关,其中渗透调节能力直接决定了贝类适合生长的环境[17]。尤仲杰等[18]研究发现,墨西哥湾扇贝幼虫和稚贝在不适的盐度下,其围心腔压力增大,心跳减慢,对外界的刺激反应迟钝等,处于这种状态下的幼虫抵抗力下降,代谢缓慢或停止,易死亡。
本研究表明:大竹蛏受精卵孵化及变态的适宜盐度为18~30,最适盐度范围22~30。这与文蛤孵化期最适盐度17.3~28.4[19]相近,与文蛤浮游幼体的最适盐度15.9~22.6[19]不同。由于大竹蛏分布在潮下带至水下20 m水深的浅海,盐度较近海高,故其对低盐的耐受能力较差;而文蛤长期生活在有淡水注入的河口附近,自然进化成对盐度的广泛适应性,并表现出对低盐的适应性强于高盐[14]。本结果与陈爱华[12]对大竹蛏稚贝的研究一致;与青蛤20~25为最适盐度(变态率在84.1%~88.0%)[15]相近,与毛蚶受精卵孵化最适盐度22~26[16],毛蚶幼虫生长最适盐度20~25相近。这也表示,大竹蛏受精卵及浮游幼虫与大多数海洋双壳贝类相同,对盐度要求较高,在适盐范围内生长发育较快。
3.3耐盐耐温实践指导意义
海水温度和盐度是影响海洋生物存活和生长的重要环境因子,当二者中的一个因子控制在安全范围,通过单因素实验能够反映生物对温度或盐度的适应性[20],这也是本试验研究的主要内容。贝类对温度和盐度的适应能力与种类的遗传性和个体的生理状态有关,而与其个体大小关系不明显,同种类的所有大小的个体都以相同方式对温度和盐度作出反应[21],这与本研究的结果一致。许多学者[6-7]认为生物体对环境的适应能力受许多环境因子的共同影响,其中温度和盐度作为养殖环境的主要因子,对双壳贝类的生长、存活和繁殖有显著影响[22-24]。有关研究表明,温度和盐度不但影响贝类的胚胎发育速度和孵化率,也影响所孵化幼虫的健康状态。当温度或盐度超过贝类胚胎发育的忍受范围时,将引起孵化率明显下降,部分能发育到幼虫的,其畸形率也明显增大[25-26]。
孵化率及变态率的高低对苗种繁育影响重大,故其幼体孵化及变态的适宜温度和盐度范围在其人工苗种生产中具有十分重要的指导意义。建议在大竹蛏苗种繁育的过程中,将温度控制在21~27℃内,盐度范围22~30内为宜。
参考文献:
[1]潘星光.缢蛏的生态观察与食性分析 [J].动物学杂志,1959,3(8):355-357.
[2]戴中杰.大竹蛏软体部营养成分分析及评价[J].集美大学学报,2002,7(4):304-308.
[3]戴中杰.大竹蛏软体部的氨基酸分析[J].莆田学院学报,2002,9(3):32-35.
[4]肖国强,柴雪良,邵艳卿,等.大竹蛏的繁殖生物学[J].海洋科学,2009,33(10):21-25.
[5]陈爱华,姚国兴,吴杨平,等.吕四渔场海区大竹蛏的繁育与生长[J].海洋渔业,2012,34(3):277-284.
[6]吴杨平,陈爱华,姚国兴,等.大竹蛏生物学特性的初步研究[J].水产科学,2011,30(12):764-767.
[7]曹琛,闫喜武,李晓冬,等.大竹蛏生产性人工育苗技术研究[J].齐鲁渔业,2012(6):4-7.
[8]王雪梅,路宜华,丰爱秀,等.大竹蛏健康苗种培育新模式的研究[J].水产养殖,2012(8):14-16.
[9]侯和要,牟乃海,宋全山,等.大竹蛏人工繁育技术研究[J].齐鲁渔业,2004,21(6):32-35.
[10]陈爱华,姚国兴,张志伟.大竹蛏生产性人工繁育实验[J].海洋渔业,2009,31(1):66-72.
[11]闫喜武,赵生旭,张澎,等.培育密度及饵料种类对大竹蛏幼虫生长、存活及变态的影响 [J].大连海洋大学学报,2010,25(5):386-390.
[12]陈爱华,张志伟,姚国兴,等.环境因子对大竹蛏稚贝生长及存活的影响 [J].上海水产大学学报,2008,17(9):559-563.
[13]陈爱华,姚国兴,张志伟,等.温度、盐度和底质对大竹蛏稚贝生长及存活的影响 [J].热带海洋学报,2010(5):94-97.
[14]侯和要,王君霞,彭作波,等.不同盐度对大竹蛏存活的影响[J].齐鲁渔业,2004,21(5):5-6.
[15]王丹,徐善良,尤仲杰,等.温度和盐度对青蛤孵化及幼虫、稚贝存活与生长变态的影响[J].水生生物学报,2005,29 (5):495-501.
[16]沈伟良,尤中杰,施祥元.温度与盐度对毛蚶受精卵孵化及幼虫生长的影响[J].海洋科学,2009,33(10):5-8.
[17]薛凌展,阙华勇,张国范,等.盐度对近江牡蛎幼虫生长及存活的影响[J].海洋科学,2007,31(9):73-76.
[18]马洪明,刘晓伟,麦康森,等.盐度突降对栉孔扇贝(Chlamys farreri)抗病力指标的影响[J].高技术通讯,2006,16 (7):746-751.
[19]陈冲,王志松,随锡林.盐度对文蛤孵化及幼体存活和生长的影响[J].海洋科学,1999(3):16-18.
[20]Castagna M.Culture of the bayscallop,Argopecten irradinans Lamark,in Vinrginia[J].Marine Fisheries Review,1975 (37):19-24.
[21]何义朝,张福绥.墨西哥湾扇贝稚贝对盐度的耐受力[J].海洋学报,1999,21(4):87-91.
[22]林笔水,吴天明.盐度和温度同缢蛏稚贝存活于生长的关系[J].水产学报,1986,10(1):41-45
[23]尤仲杰,徐善良,边平江,等.海水温度和盐度对泥蚶幼虫和稚贝生长及存活的影响 [J].海洋学报,2001,23(6):108-113.
[24]刘志刚,王辉,栗志民,等.墨西哥湾扇贝高起始致死温度的研究[J].中国水产科学,2007,14(5):778-785.
[25]何义朝,张福绥.盐度对海湾扇贝不同发育阶段的影响[J].海洋与湖沼,1990,21(3):197-203.
[26]何义朝,张福绥.温度对贻贝胚胎发育的影响[A].贝类学论文集(第一集)[C].北京:科学出版社,1983:133-142.
资助项目:农业科技成果转化资金项目(2014GB000363)
doi:10.3969/j.issn.1004-2091.2016.08.006
收稿日期:(2016-01-28)
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
当代水产(2021年5期)2021-07-21
生物学通报(2021年5期)2021-03-16
海洋通报(2020年3期)2020-10-20
科普童话·百科探秘(2020年3期)2020-05-11
启迪与智慧·下旬刊(2019年9期)2019-01-19
当代县域经济(2018年9期)2018-09-05
文苑·经典美文(2017年6期)2017-06-12
广西科学院学报(2016年4期)2017-01-07
当代县域经济(2015年12期)2015-03-20
