
黄海近岸潮汐锋海域蟹类幼体日间垂直迁移特征❋
2022-09-05 04:33叶振江彭玉强何天庆何青松张文超张艺笑
中国海洋大学学报(自然科学版) 2022年9期
叶振江, 彭玉强, 何天庆, 何青松, 张文超, 张艺笑
(中国海洋大学水产学院, 山东 青岛 266003)
海洋鱼类、蟹类的早期生活史对群体资源量影响巨大,长期以来一直是国内外渔业资源研究的热点[1-6]。蟹类的早期生活史涉及亲蟹繁殖、幼体发育、生长、死亡、栖息地选择和幼体输运等诸多方面。国外对海洋蟹类早期生活史的研究相对较多,研究表明不少蟹类幼体存在昼夜垂直迁移特性[7-9]。在早期补充过程中,蟹类幼体的垂直迁移是关键的生态现象,它与个体摄食、幼体向育幼场的输运与扩散、栖息地选择和补充成功率变动等密切相关[10-12]。对蓝蟹(Callinectessapidus)的研究表明,其幼体不同阶段呈现复杂的垂直迁移模式,并对幼体输运及早期补充产生影响。在游泳能力弱的溞状幼体阶段,个体通过昼夜垂直迁移模式选择水层,从而借助潮流进行水平方向的迁移,从河口附近区域移向大陆架开放水域;随着个体的发育,其晚期溞状幼体和大眼幼体对水压变化的响应特征与早期溞状幼体显著不同,垂直迁移与分布特性也发生改变,稚蟹的补充在大眼幼体向河口区移动的过程中完成,表层流控制着沿岸区域所有蓝蟹幼体的输运,幼体输运是否成功取决于幼体生活史的过程与海洋学事件是否相匹配[13]。蟹类幼体的垂直迁移随着不同物种的生物学属性及不同的栖息生境,可呈现为日间或潮周期不同类型的时间模式[14-15],而内在的生物钟、外部环境变动周期性刺激或二者的结合是引起这些幼体周期性垂直迁移活动的原因[16-18]。
记述中国的海洋蟹类约1 073种[19],其中黄海中部海域分布的蟹类约90余种,其中不乏三疣梭子蟹(Portunustrituberculatus)、日本蟳(Charybdisjapo-nica)这些重要经济种[20-21]。限于海洋蟹类幼体形态发育记述严重不全,仅有三疣梭子蟹、日本蟳、天津厚蟹(Helicetientsinensis)等少数零星报道[22-24],国内关于海洋蟹类幼体垂直迁移特性在内的早期生活史的系统研究基本处于空白,至今仅对长江流域中华绒螯蟹(Eriocheirsinensis)早期补充有过深入报道[25-27],这种情况不利于蟹类资源的科学保护、合理开发与可持续利用。
黄海中部是渔业资源动物的重要产卵场和育幼场,有显著的潮汐锋水文结构[28]。本文以贯穿区域潮汐锋的35°N断面为调查区域,进行夏季蟹类幼体资源的综合断面调查,并在潮汐锋核心区及内外侧分别设连续观测站,进行环境与蟹类幼体24 h垂直分布的连续采样,旨在阐明蟹类幼体在不同水文结构背景下的垂直迁移规律,探索垂直迁移对幼体输运过程的影响,为深入开展蟹类资源早期生活史研究、科学保护和开发蟹类资源提供参考。
1 材料与方法
1.1 调查航次与站位
实验于2020年7月24日—30日进行。于黄海中部35°N断面设置17个调查站位(#1~#17),各站位间距为5 n mile。其中#1、#6和#17为3个连续站观测站位(#1:119°54.829′E,35°01.474′N;#6:120°24.151′E,35°00.090′N;#17:121°31.000′E,35°00.090′N),其余站位为一般断面观测站位,均匀分布于3个连续观测站点之间,沿35°N线构成一条水平观测线,经度跨越119°54.829′E—121°31.000′E。在连续站站位进行24 h昼夜蟹类幼体丰度与环境观测,每2 h观测一次,24 h内共观测12次,每次首先进行CTD(型号:德国Sea & Sun Technology公司CTD-75M型)环境观测,获取温度、盐度的垂直剖面数据,然后进行蟹类幼体水平和垂直拖网采样,并根据温度剖面特征将水体分为上、下两层,分别进行蟹类幼体垂直拖网采集。#1、#6和#17站上下层分界分别为15、15和20 m,其余站位于航行过程中进行海洋环境的CTD观测和蟹类幼体样品的水平拖网采集。生物样品采集使用大型浮游生物网(网口直径80 cm,网目孔径0.505 mm,网衣长2.8 m)进行,网口系流量计校正滤水量,每个水平站位以3 kn左右船速拖网10 min,垂直生物样品采集则在网口上附上铅锤进行。垂直分层样品采集时利用附加绳索控制网口开闭。同时,网口上附小型温、深记录探头,用于记录网具采样水深。

表1 连续站蟹类幼体样品组成
1.2 样品处理
采集生物样品后,现场马上利用酒精固定样品,并带回实验室,使用解剖镜挑出蟹类幼体样品,根据形态进行初步分类,分别记录溞状幼体和大眼幼体的数量,用于后续分析,挑选过程中根据幼体较高的的额棘长度及与身体比例来排除可能混入的歪尾目幼体。由于中国海域蟹类幼体的形态发育报道甚少,仅有少数物种的记述,物种形态鉴定较困难,因此并未对蟹类幼体进行物种鉴定,仅分析了不同阶段幼体数量的分布。部分出现频率高的溞状幼体和大眼幼体的照片见图1。

((a)~(f):溞状幼体,(A)~(F):大眼幼体。(a)~(f):Zoea;(A)~(F):Megalopae.)
1.3 数据分析
同一站位由于采样方式、调查时段、滤水量及生物分布的不同,水平、垂直及上、下水层获得的样品数量绝对值存在差异。为方便分析,作者采用相对丰度指数K作为表征蟹类幼体垂直分布数量的丰度指标。其定义为:
K=Ni/N。
式中:K为相对丰度指数,代表该次采样生物量密度占该站位12次采样总生物量密度的相对比例;Ni为连续站第i次采样获得的生物量密度(ind./m3);N为连续站该采样方式昼夜12次采样获得的生物量密度之和。
采用断面上1~17站位的常规垂直采样数据进行幼体丰度的水平分布趋势分析。为提高数据代表性,使用3个连续站所有的幼体采集数量分析大眼幼体与溞状幼体相对比例的空间变化。
2 结果
2.1 调查海域的物理海洋环境
#1站位垂直剖面水温在22.71~24.04 ℃之间变化,在19:00和9:00左右出现低温水舌的明显上涌,在15:00和4:00左右出现高温水舌的下探(见图2a)。#6站位垂直剖面水温在21.21~24.14 ℃之间变化,有明显的潮汐锋核心区上下水层混和现象,较高水温在15:00—23:00于水深5 m以上水层出现,在21:00和11:00左右有微弱的低温水舌上涌,在5:00左右有微弱高温水舌下探(见图2b)。#17站位受黄海冷水团势力影响,水温垂直分布变化最大,变化范围为14.83~22.99 ℃,显著的温度跃层位于20 m水深左右,在15:00、21:00和11:00左右出现低温水舌上涌现象,但止于温度跃层下方;在18:00和5:00左右出现表层高温水舌下探(见图2c)。
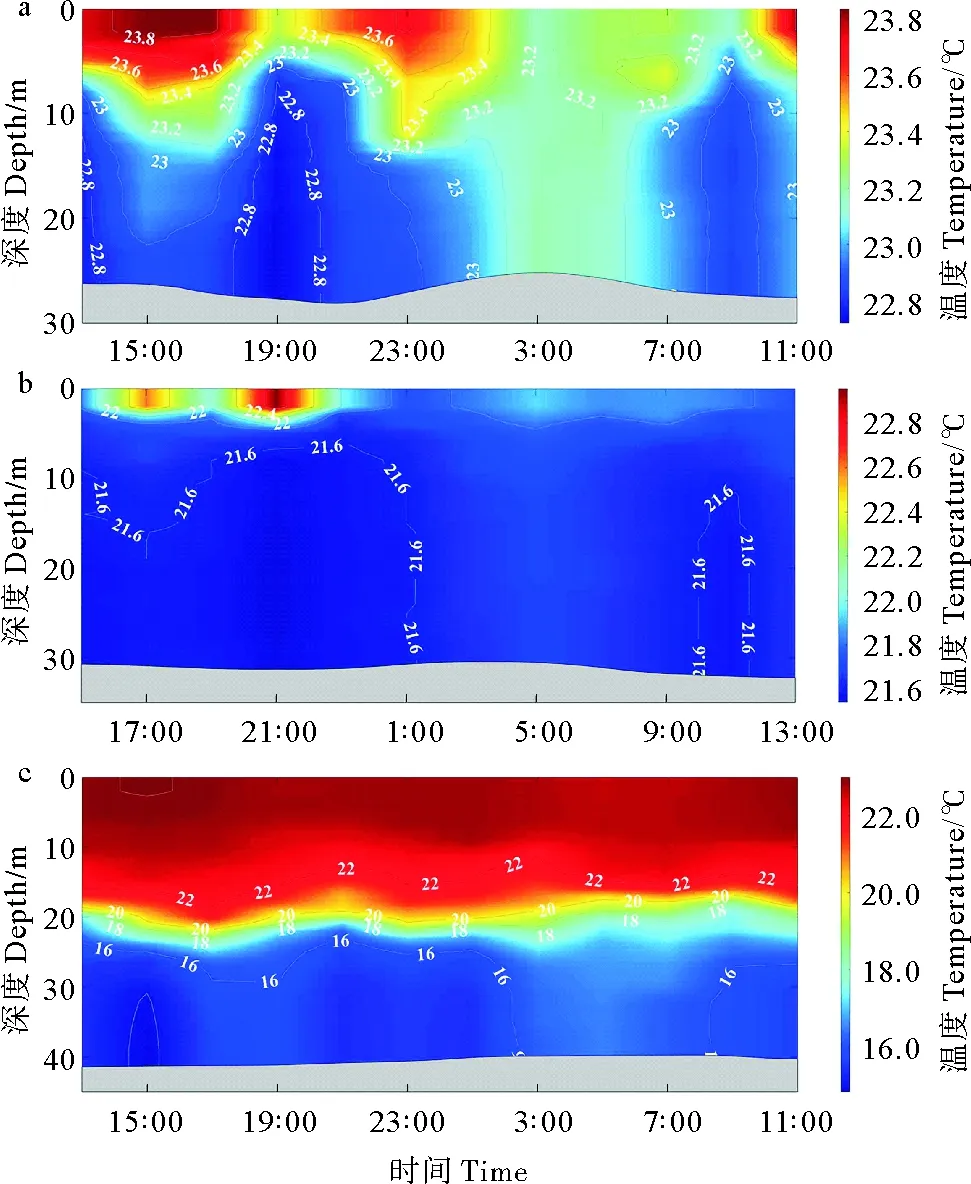
(a: #1站;b: 6#站;c: #17站。a: Station #1; b: Station #6; c: Station #17.)
#1、#6和#17站位的盐度变化范围分别为28.67~30.75,31.13~31.38和30.89~32.41,各站位皆有显著的盐度垂直结构,盐度垂直结构随时间的变化与随温度的变化相类似(见图3)。其中,#1站位整体盐度较低,#6和#17站位盐度依次升高,#17站位在22~25 m左右水深出现盐度跃层(见图3)。
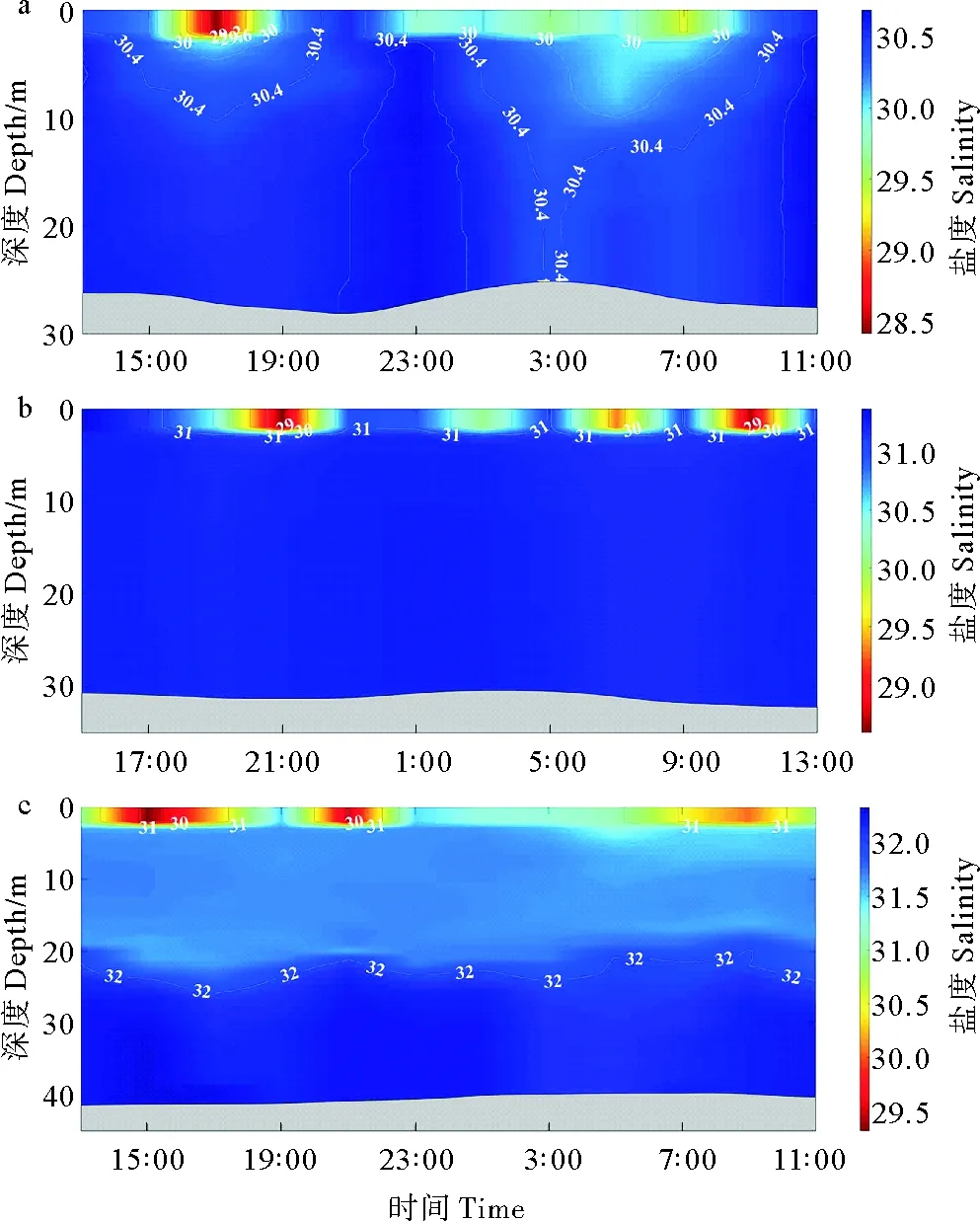
(a: #1站;b: 6#站;c: #17站。a: Station #1; b: Station #6; c: Station #17.)
2.2 蟹类幼体的水平分布
沿调查断面的蟹类幼体水平分布呈从近岸向远岸逐渐减少的分布格局(见图4)。3个连续观测站中,#1、#6和#17站分别采集蟹类溞状幼体66 897、46 115和21 671尾,采集大眼幼体1 104、2 467和1 486尾。同时,大眼幼体所占比例#1站<#6站<#17站。
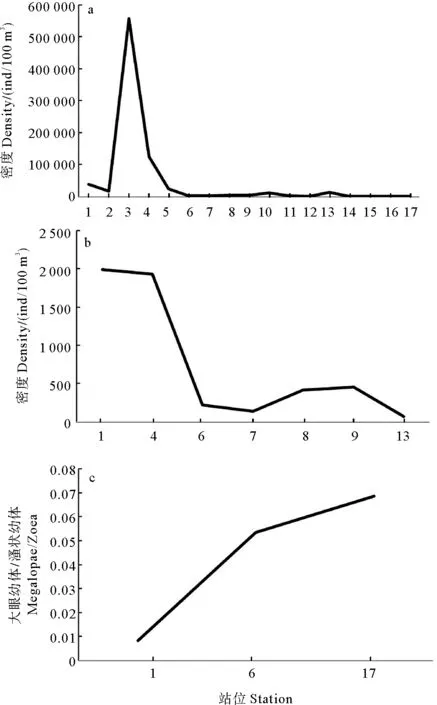
(a: 溞状幼体;b: 大眼幼体;c: 大眼幼体/溞状幼体。a: Zoea;b: Megalopae;c: Megalopae/zoea.)
2.3 蟹类幼体垂直分布
2.3.1溞状幼体 溞状幼体数量垂直分布呈现显著的昼夜变化(见图5)。
从水平网采数量的时间分布来看,#1站数量峰值出现于3:00和21:00,相对丰度指数分别为0.31和0.20;#6站数量峰值出现于23:00和3:00,相对丰度指数分别为0.39和0.21;#17站数量峰值出现于21:00和3:00,相对丰度指数分别为0.35和0.31(见图5)。
垂直网采溞状幼体数量亦存在两个峰值,但峰值时段较水平丰度推迟2 h左右。#1站溞状幼体主要出现于23:00—7:00,数量峰值出现于23:00和7:00,相对丰度指数分别为0.27和0.29;#6站溞状幼体主要出现于23:00—7:00,数量峰值出现于23:00和5:00,相对丰度指数分别为0.77和0.09;#17站溞状幼体主要出现于21:00—5:00,数量峰值出现于23:00和5:00,相对丰度指数分别为0.37和0.31(见图5)。
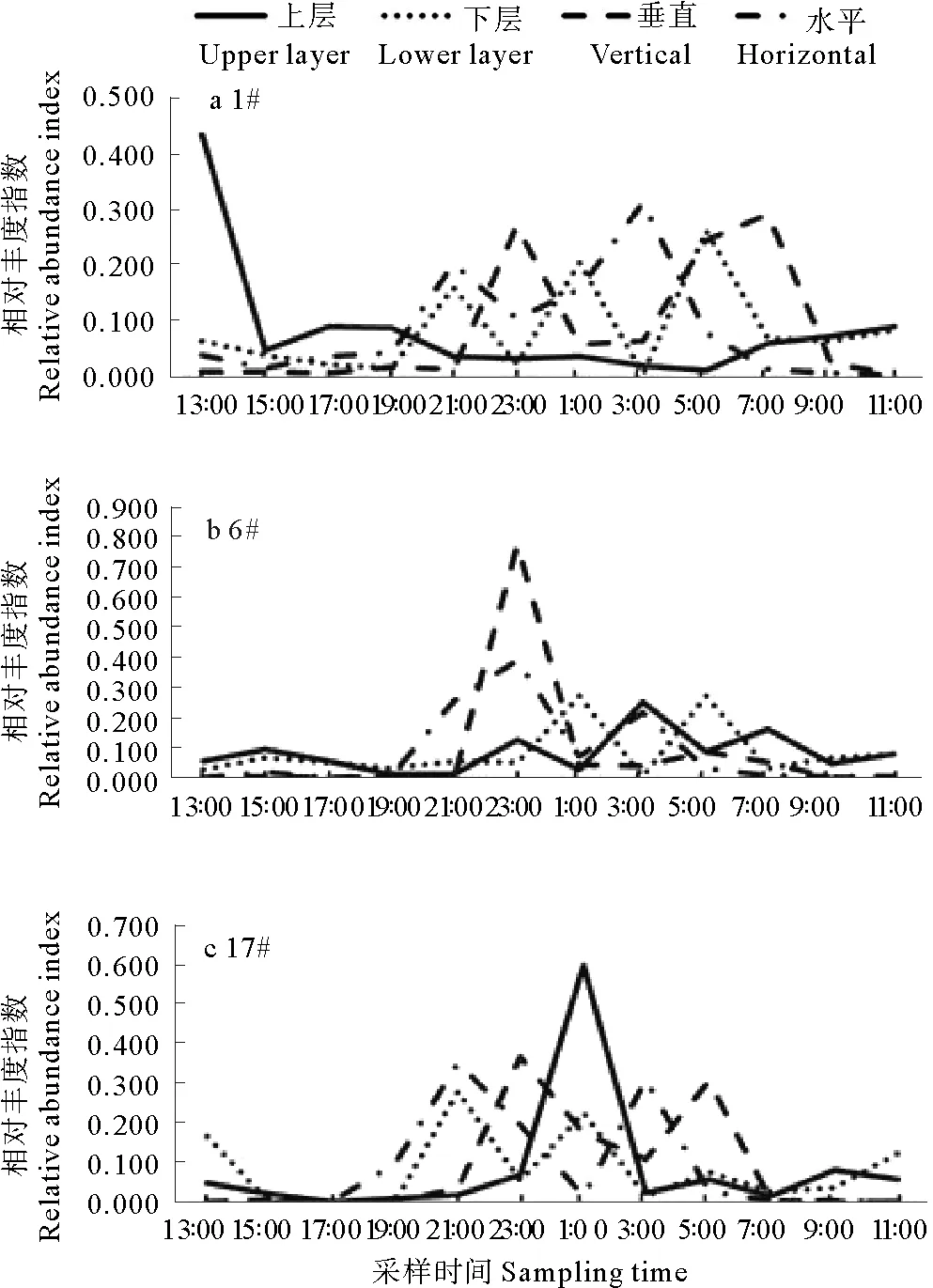
图5 蟹类溞状幼体垂直分布的日间模式
上、下水层网采数据显示,3个站位下层数量皆以夜间为多,峰值一般出现在21:00、1:00和5:00;上层数量中,#1站最高值出现于13:00,最低值出现于5:00,其它2个站位的数量峰值皆以夜间较多,有1~3个数量峰值。
2.3.2 大眼幼体 大眼幼体数量垂直分布亦呈显著的昼夜变化(见图6)。水平网采结果显示,大眼幼体在#1站主要出现于21:00—5:00,数量高峰出现于3:00—5:00;在#6站主要出现于21:00—3:00,数量高峰出现于23:00—1:00;在#17站主要出现于21:00—3:00,数量高峰出现于3:00和23:00。除上述主要时段外,其它时段水体表层鲜有大眼幼体分布(见图6)。
垂直网采数量亦呈现显著的昼夜变化。大眼幼体在#1站主要出现于23:00—7:00;在#6站主要出现于23:00—7:00,数量高峰出现于23:00—3:00;在#17站主要出现于1:00—3:00(见图6)。上、下水层大眼幼体数量,#1站上层数量峰值出现于13:00和7:00,下层主要分布于夜间,数量峰值出现于1:00和5:00;#6站上层大眼幼体主要出现于夜间,数量峰值出现于23:00、3:00和9:00,下层大眼幼体主要出现于凌晨到上午,数量峰值出现于5:00和11:00;#17站上层大眼幼体主要出现于夜间,数量峰值出现于1:00和5:00点,下层大眼幼体出现于凌晨1:00到上午9:00点,数量峰值出现于1:00—3:00和9:00(见图6)。
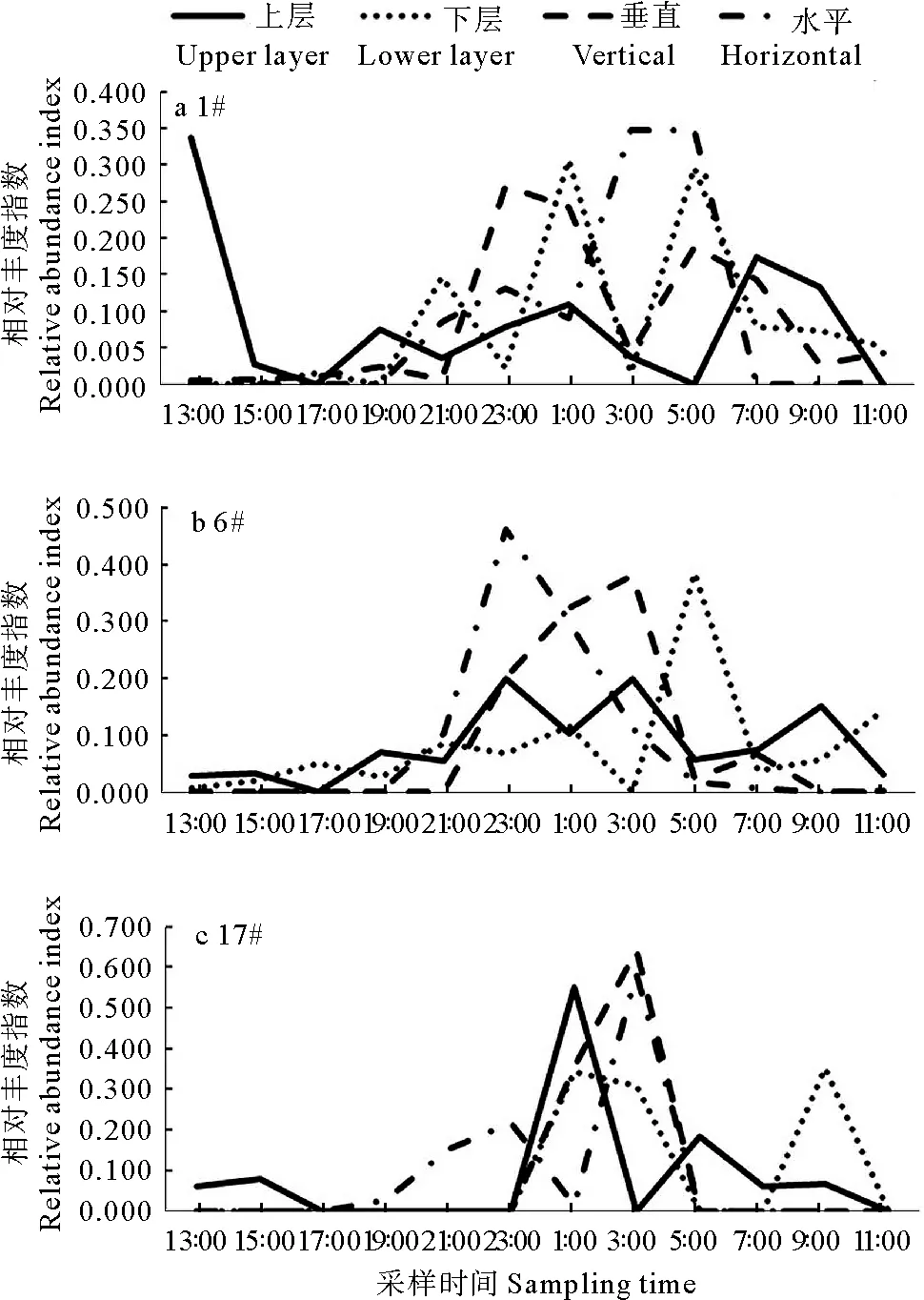
图6 蟹类大眼幼体垂直分布的日间模式
3 讨论
3.1 蟹类幼体的垂直迁移模式
相关研究发现,很多蟹类幼体具有显著的垂直迁移现象[7-9]。蟹类幼体一般游泳能力尚较弱,如蓝蟹溞状幼体游泳速度只为0.5~2.0 cm/s,这个速度尚不能与水平方向的海流抗衡和通过自主游泳实现水平输运,但是幼体可通过昼夜的垂直迁移来选择不同的水层,从而实现定向水平输运的目的。对于不同蟹类,不仅繁殖时间可能不同,其幼体的垂直分布特性亦会存在差异,并在幼体输运去向上产生差异[7]。因此,蟹类幼体的垂直迁移不仅是摄食的需要,亦在幼体扩散过程中起着重要作用。
本文结果中,溞状幼体夜晚上浮、白天沉底的日间运动模式显著。从垂直采样结果观察,#1和#6站蟹类溞状幼体从9:00始即几乎全部沉入水底,21:00后开始上浮;#17站则7:00沉于水底,19:00后开始上浮。与溞状幼体相比较,大眼幼体在各站位出现的时段更为集中,特别是#6和#17站,白天大量时段垂直与水平采样没有幼体分布,即使是#1站,虽然垂直采样数量在日间模式上有所分散,但水平采样在7:00—19:00之间几没有任何分布,这表明大眼幼体的垂直分布水层较溞状幼体更为偏下,同时,作者认为这是大眼幼体运动能力增强后对环境的自主选择能力增强所致。
3.2 幼体垂直迁移的环境影响机制
相关研究表明,一些环境因素包括温度、盐度、水压、重力、潮汐、光周期等外部环境因素在蟹类幼体的垂直迁移和水平输运过程中起着重要作用[14]。同时,单细胞藻类、轮虫无节幼体是蟹类幼体的主要饵料生物,各类浮游动物亦存在显著的昼夜垂直移动现象,这可能也是引起蟹类幼体垂直迁移的重要因素[29-31]。本调查海域为半日潮海区,潮时每天较前一天有半个多小时的延迟(见图7),从#1到#17站历时5 d,潮时有显著的改变,但3个站位的幼体垂直分布日间模式并未有相应的较大调整,因此潮水涨落并非本区域蟹类幼体垂直迁移的直接物理触发因素。在日照、潮流、地形等综合因素作用下,3个连续观测站的温度、盐度剖面分布呈显著的日间变化。在每个站位水体表层的溞状幼体达到数量峰值时所对应的环境温度与盐度并非固定值,特别在最外侧的#17站,该站受黄海冷水团影响,在20 m左右存在明显的温度跃层和盐度跃层,环境垂直剖面的温、盐度变动范围可高达14.83~22.99 ℃和30.89~32.41,但溞状幼体的垂直迁移节律并未受到明显影响,这表明幼体在垂直迁移过程中具有较强的适应温、盐变化的能力。调查期间日出时间稳定在5:06左右,光周期可能是影响幼体垂直迁移的主要环境刺激因素。同时,溞状幼体在3个连续站的垂直分布与水平分布皆在夜晚出现2个峰值,这是由于不同物种或不同发育阶段幼体垂直迁移特性不同所致,还是存在其它原因,有待于今后进一步研究。

(仿自https://www.cnss.com.cn/tide/。Imitated from https://www.cnss.com.cn/tide/.)
3.3 幼体输运趋势
幼体丰度从近岸向远岸逐渐减少,大眼幼体与溞状幼体的相对比例同时增大。大眼幼体的游泳速度可达5 cm/s,这个速度在局部水平移动如附底过程中可能会发挥重要作用,但尚不足以完全支持自主的水平输运[32]。离岸方向大眼幼体数量的增加可能是蟹类幼体以个体垂直迁移方式借助潮流,从近岸向远岸输运的结果。然而,蟹类幼体的栖息地选择与输运机制相当复杂,不同种或发育阶段的幼体对潮汐的响应和日间分布模式有可能不同[33-34]。随着蟹类溞状幼体的发育其垂直游泳速度提升,也会对溞状幼体的垂直移动产生一定影响;大眼幼体的垂直游泳能力显著高于溞状幼体,其选择垂直分布的水层深度也就更为宽泛,垂直迁移模式也可能发生显著改变[13]。因而,关于蟹类幼体的水平输运与育幼场分布,有待于在查明各物种及不同发育阶段的日间垂直分布模式的基础上,运用水动力学模型开展数值模拟,并经过更多的调查研究来验证。同时,由于目前水平采样过程主要在白天进行,在幼体存在显著昼夜垂直迁移的背景下,除了3个连续观测站位外,其它14个平面采样站位的数据在幼体丰度上存在较大局限性。今后宜将蟹类幼体调查安排在夜间进行,并使用国际上通用的Bongo网进行双斜拖采样,以保障调查效果。
4 结语
本文研究结果表明,黄海蟹类幼体具有显著的日间垂直迁移现象,呈现夜晚上浮、白天下沉的特征;夜晚幼体在表层出现的峰值,在垂直方向的分布大眼幼体较溞状幼体更为偏下;离岸较远的站位大眼幼体的数量相对更高。在物种分子生物学分类基础上建立不同蟹类幼体形态学分类标准,探索不同蟹类幼体的日间垂直迁移模式的差异,研究该差异对幼体的输运、栖息地分布及早期生活史的作用机制,将是今后的重点研究方向。
猜你喜欢
河北渔业(2022年10期)2022-10-15
文萃报·周二版(2022年10期)2022-03-19
水上消防(2021年3期)2021-08-21
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
河北渔业(2018年4期)2018-05-08
安徽农学通报(2016年24期)2017-01-12
河北渔业(2015年1期)2015-01-19
江苏农业科学(2014年8期)2014-10-23
