
Permeabilization of Escherichia coli with ampicillin for a whole cell biocatalyst with enhanced glutamate decarboxylase activity☆
2016-05-30 12:54WeiruiZhaoShengHuJunHuangPiyuKeShanjingYaoYinlinLeiLeheMeiJinboWang
Weirui Zhao ,Sheng Hu ,Jun Huang ,Piyu Ke ,Shanjing Yao ,Yinlin Lei,Lehe Mei,*,Jinbo Wang
1 School of Biotechnology and Chemical Engineering,Ningbo Institute of Technology,Zhejiang University,Ningbo 315100,China
2 Department of Chemical and Biological Engineering,Zhejiang University,Hangzhou 310027,China
3 School of Biological and Chemical Engineering,Zhejiang University of Science and Technology,Hangzhou 310023,China
1.Introduction
γ-Aminobutyric acid(GABA)is a non-protein amino acid thatacts as an inhibitory neurotransmitter in animals and directly responds to stress[1].It exhibits well-known physiological functions in animals and humans including the induction of hypotensive,diuretic,and tranquilizing effects[2–4].GABA is also a strong secretory analog of insulin and can prevent diabetes[5].Recently,GABA has been used as a component of pharmaceuticals and functional foods.In addition,GABA shows promising applications as a novel building block for the environmentally safe synthesis of multitudinous nitrogen-containing industrial chemicals including N-methylpyrrolidone[6]and biodegradable plastics such as polyamide 4[7,8].Because of its significance in the food,pharmaceutical,and chemical industries,various chemical or biological methods for GABA preparation have been developed.In contrast to chemical synthesis methods,GABA biosynthesis techniques are promising due to their simple reaction procedures,environmental compatibility,and mild reaction conditions[9,10].Glutamate decarboxylaxe(GAD)is a unique enzyme known to catalyze irreversible α-decarboxylation of L-glutamate or its salts to GABA[11].Therefore,numerous studies have been conducted to develop biocatalysts with high GAD-specific activities[7,12–15].
In biotransformation procedures,whole-cell systems are usually preferred over isolated enzymes because these eliminate the requirementfor tedious and expensive enzyme isolation/purification processes[16,17].Therefore,much attention has been given to developing wholecell biocatalysts with high cell-associated GAD activity for commercial use.Forthis purpose,recombinant GADs have been expressed in various species to improve GABA production[7,12–14].Although overexpression of GAD is an effective means to generate cell preparations with high enzyme activity,the reaction rates and yields of this particular synthesis method using whole-cell biocatalysts are strongly limited by the cell envelope,which acts as a barrier against the diffusion of substrates(L-Glu)and products(GABA)[13].
Therefore,some measures should be taken to improve cell permeability and enhance the GABA synthetic capability of whole-cell biocatalysts[13].A common way to improve cell permeability is by treating cultivated cells with chemical permeabilizers such as cetyltrimethyl ammonium bromide(CTAB)[18],Triton X-100[19],toluene[20],or EDTA[21]as well as a variety of physical methods[17,22].Although such treatments improve cell permeability,these also require severaladditionaland tedious processing steps thatare especially undesirable for large-scale production[16].For example,permeabilizing cultivated cells requires the following steps:(1)harvesting the cultivated cells by centrifugation;(2)treating the cells with permeabilizers;(3)recovering the cells by centrifugation;and(4)washing the cells to remove the residual permeabilizers[19].Therefore,developing permeabilization methods without the extra cell-permeabilization steps will be bene ficial for large-scale GABA production using wholecell biocatalysts.
Cell wall-synthesis inhibitors suppress cell wall formation,and surfactants modify cell envelope structures during cell cultivation[23,24].Thus,itis possible to use these to improve cellpermeability without the extra cell-permeabilization steps.The recombinant Escherichia coli strain BL21(DE3)-pET28a-gadB over-expressing GAD from Lactobacillus brevis CGMCC 1306 was prepared in our laboratory and used as a candidate for GABA production[25].We investigated the utility of cell-wall synthesis inhibitors and surfactants as tools to improve its cell permeability and enhance cell-bound GAD activity.This enhanced the GABA synthetic ability of BL21(DE3)-pET28a-gadB cells without extra cell-permeabilization steps after cell cultivation.
2.Materials and Methods
2.1.Chemicals
A GABA standard was purchased from Acros Organics(Geel,Belgium).Dansyl chloride(Dns-cl)was obtained from Tokyo Chemical Industry Co.,Ltd.(Japan).PVA,CTAB,sodium dodecyl sulfate(SDS),Tween 80,and Span 80 were purchased from China Medicine Co.Ltd.(China).Isopropyl-β-D-thiogalactoside(IPTG),5′-pyridoxal phosphate(PLP),kanamycin,and ampicillin were provided by Shanghai Sangon Co.,Ltd.(China).Unless specified,all other chemicals were of analytical grade or higher.The BL21(DE3)-pET28a-gadB used here was constructed and stored in our laboratory[25].
2.2.Strain and culture conditions
E.coli BL21(DE3)-pET28a-gadB cells were cultivated in 100 ml of LB-media containing 50 μg·ml−1kanamycin at 37 °C and constant shaking at200 r·min−1.The IPTGwas added to the cultures ata finalconcentration of 0.5 μmol·L−1to induce GAD expression after the bacteria reached an opticaldensity of0.65–0.75(600 nm).In parallel,ampicillin,CTAB,SDS,Tween 80,or Span 80 were added to different cultures to study its effects on GAD activity.The cells were then cultured at 30°C with shaking at 150 r·min−1for an additional 6 h.
2.3.Preparation of cell free extracts
Total cell lysates from ampicillin-permeabilized cells and control cells were prepared according to the method described by Fan et al.[25].Brie fly,cells were collected by centrifugation(12000 g,4°C,10 min),suspended in disruption buffer(0.1 mol·L−1phosphate buffer,1 mmol·L−1phenylmethanesulfonyl fluoride,pH 7.5),and then sonicated at 4°C(Sonifier JY92;Xinzhi Biotechnology Institute,China;output of 300 W,duty time of 3 s,interval time of 6 s,for 90 cycles).Celldebris wasremoved by centrifugation at12000 g at4°C,and the supernatants were used as cell free extracts.
2.4.Immobilization of permeabilized cells
Immobilization of the permeabilized cells was performed according to the method described by Liu et al.[26].Brie fly,8.0 g PVA and 1.0 g sodium alginate(SA)were dissolved in 100 ml boiled distilled water.Once the solution cooled down to 40°C,400 mg of cells(dry cell mass,DCM)were added to the solution and incubated for 5 h.The solution was then mixed thoroughly to yield a homogeneous cell/PVA-SA suspension.The cell/PVA-SA solution was then added to a curing agent(6%boric acid and 1%CaCl2solution)using a syringe pump at a rate of 0.32 ml·min−1,and the beads were incubated in this curing agent at 30°C for 8 h until solidification was complete.The sample was then washed with distilled water to remove the excess boric acid.
2.5.GAD activity assays
GAD activity was measured by determining the amount of GABA formed at 37 °C in a reaction mixture containing 0.2 mmol·L−1sodium acetate buffer(pH 4.8),75 mmol·L−1sodium L-glutamate(L-MSG),and 0.1 mmol·L−1PLP.The GABA concentration was assayed by using HPLC as described by Huang et al.[10].One unit(U)of GAD activity was defined as the amount of catalyst that produces 1 μmol GABA per minute under the given assay conditions[13].Specific activity was defined as U·mg−1(DCM)cells.
The total cell-bound GAD activity(TGA)per ml of culture medium was the main criteria for agent selection and process optimization.This was selected because the additive agents might affect the total cell-bound GAD activity per volume of culture medium in terms of cell-bound GAD activity of BL21(DE3)-pET28a-gadB and its cell biomass.The total cell-bound GAD activity of 1 ml of culture(U)was equal to the cell-bound GAD activity of the whole cells(U·mg−1)×the the DCM in a 1-ml culture(mg).
2.6.Morphology studies
The E.coli cells were harvested by centrifugation at 2000 g for 10 min,and the pellets were fixed in 2.5%glutaraldehyde at 4°C overnight.Subsequently,the cell pellets were washed 3 times with phosphate buffer(0.1 mol·L−1,pH 7.0)and transferred to 1%osmic acid for 1 h.The pellets were washed again in phosphate buffer and dehydrated in graded ethanol series(50%–100%).Next,the samples were treated with isoamyl acetate and washed twice(15 min).Finally,the samples were dried in a critical point drier,sputter-coated with gold–palladium,and viewed under an environmentalscanning electron microscope(SEM,Philips XL-30 ESEM).
For transmission electron microscopy(TEM)studies,the E.coli cells was collected, fixed,and dehydrated as described above for the SEM analysis.Samples were then embedded in Spurr's resin,and ultrathin sections(70 nm)were cut and stained with uranyl acetate and lead citrate.These sections were observed under a JEM-1230 electron microscope(JEOL)operating at 80 kV.
3.Results and Discussion
3.1.Permeabilization of BL21(DE3)-pET28a-gadB with different additives
The cell-bound GAD activity of BL21(DE3)-pET28a-gadB cells cultured with different additives are shown in Table 1.The cell wallsynthesis inhibitor ampicillin and the cationic surfactant CTAB had inhibitory effects on E.coli cell proliferation,whereas these improved cell-bound GAD activities.Although we observed a decrease in cell biomass,the TGA/ml of the culture medium showed an overall enhancementafterthe addition ofboth ampicillin and CTAB.These findings suggest that the use of these two agents to enhance the cell-bound GAD activity in bacterial cells has practical implications.Furthermore,these two agents are cheap and widely available.Ampicillin was moreeffective than CTAB with a 3.64-fold increase in TGA at 5 μg·mL−1ampicillin.In a previous study,treating harvested E.coil GADK10 cells at 53°C for 1 h was considered the best option for permeabilization.It enhanced the cell-bound GAD activity~3-fold[13].Compared to previous results,treating E.coli cells with ampicillin resulted in comparable effects.There was no need for the extra permeabilization steps after cell cultivation.Although the anionic surfactant SDS,the non-ionic surfactant Tween-80,and Span-80 could increase cell biomass production,they did not affect cell-bound GAD activity or improve TGA at the specific concentrations utilized in the present study.Therefore,ampicillin was further evaluated as a chemical tool for the preparation of permeabilized E.coli cells.
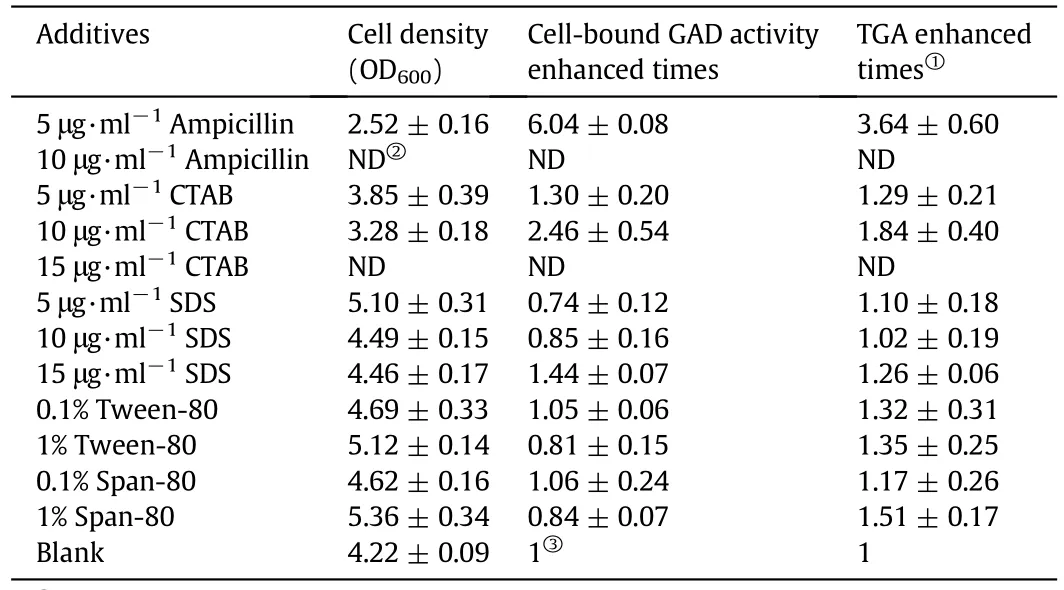
Table 1 Cell-bound GAD activity of BL21(DE3)-pET28a-gadB cells with different additives
3.2.Effect of ampicillin concentration
The effectofampicillin concentration on cellbiomass and cell-bound GAD activities in BL21(DE3)-pET28a-gadB cells is shown in Fig.1.Cell biomass decreased with increasing ampicillin concentrations.When the ampicillin concentration reached 7 μg·ml−1,cell growth was completely inhibited.A 6.04-fold relative increase in cell-bound GAD activity was achieved with 5 μg·ml−1ampicillin.However,further increases in the ampicillin concentration led to decreases in cell-bound GAD activity.The reduction in cell-bound GAD activity using low ampicillin concentrations suggests that the amount of agent was insufficient to effectively disrupt the barrier effectofthe cell envelope.The decrease in cell-bound GAD activities at higher ampicillin concentrations may be attributed to the severe decline in enzyme synthesis that resulted from disturbances in cell metabolism,which were caused by the excessive destruction of the cell envelope.
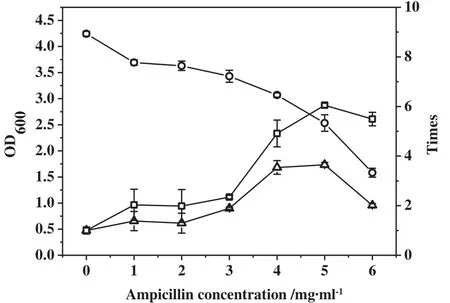
Fig.1.Effect of ampicillin concentration on cell biomass and cell-bound GAD activity of cells.(○)OD600;(□)cell-bound GAD activity enhancement;(△)TGA enhancement.
TGA also reached a maximum when the E.coli cells were cultured in 5 μg·ml−1ampicillin.TGA was enhanced by 3.64-fold and reached 2.26 U·ml−1,which was the best trade-off between the increase in cell-bound GAD activity and the decline in cell biomass.Therefore,the optimal ampicillin concentration for preparing permeabilized cells was 5 μg·ml−1.
3.3.Cell morphology of permeabilized cells
We used SEM and TEM to examine the morphological changes of E.coli cells in the presence of 5 μg·ml−1ampicillin.The SEM images showed that short and rigid rods characterized the exterior features of the control cells,whereas cells cultured in the presence of 5 μg·ml−1ampicillin were elongated and less rigid(Fig.2).Assessment of the morphology of the treated cells indicated weakening of the cell wall structure.The external morphological changes in the cells incubated with 5 μg·ml−1ampicillin were confirmed by TEM(Fig.3).However,no distinct differences in the internal structures of the E.coli cells exposed to 5 μg·ml−1ampicillin and the control cells were observed(Fig.3).Both cell populations showed uniformly distributed cytochylema and electron density,which indicated that exposure to 5 μg·ml−1ampicillin had no obvious in fluence on the internal structure of the BL21(DE3)-pET28a-gadB cells.
3.4.Effect of ampicillin treatment on GAD expression
To test whether 5 μg·ml−1ampicillin treatment in fluenced GAD expression,the expression level and soluble fraction of GAD in the BL21(DE3)-pET28a-gadB cells were determined.We could not identify any differences between 5 μg·ml−1permeabilized cells and untreated cells in terms of expression level and soluble fraction portion based on SDS-PAGE analysis(data not shown).
We next studied the total GAD activities of the cell lysates.The results showed that the total cell lysate GAD activity of 5 μg·ml−1ampicillin-treated cells[(9.15± 0.30)U·mg−1]was only 11.2%lower than that observed in the control cells[(9.69± 0.86)U·mg−1].This finding indicated that treatment with 5 μg·ml−1ampicillin had no substantial effects on GAD expression levels and GAD activity in vivo.Therefore,these results,together with findings of the morphology studies suggest that the enhancementof whole cell-bound GAD activity in 5 μg·ml−1ampicillin-treated cells was mainly due to improvements in cell membrane permeability.
3.5.Immobilization of ampicillin-permeabilized cells
To simplify the product purification process and enhance long-term operation stability,ampicillin-permeabilized BL21(DE3)-pET28a-gadB cells were entrapped in a PVA-SA-boric acid polymer.PVA increases the bead strength and durability,whereas calcium alginate improves their surface properties and reduces their tendency to agglomerate during gel-bead formation[27].The PVA-SA-boric acid beads can also minimize the negative effects of diffusion restriction in contrast to Caalginate beads because it forms open micropore structures inside the gel[26,28].

Fig.2.Scanning electron micrographs of the recombinant E.coli.(a)Control(untreated cells);(b)cells treated with 5 μg·ml−1 ampicillin.
Therefore,the immobilization ofpermeabilized BL21(DE3)-pET28agadB cells in PVA-SA-boric acid beads can potentially reduce the diffusion restriction caused by both cell envelopes and immobilized materials.The particles of immobilized cell were sphere,with a diameter of 2–3 mm.The operation stability of immobilized,permeabilized beads generated in the present study was investigated in terms of its reusability in the biotransformation of L-MSG to GABA at 37°C(Fig.4).The results showed that the GABA yield with immobilized and permeabilized cells retained 46%of the original yield afterrecycling the immobilized cells 10 times.In contrast to prior studies,the activity of the immobilized E.coli GAD in the sodium alginate and carrageenan gel remained at 50%during the 10th use[9]when operated at 37°C.The ampicillin-permeabilized cells immobilized in PVA-SA-boric acid beads also showed comparable stability and enabled efficient GABA production.Storage stability is another critical factor for the application of biocatalysts.In the present study,the immobilized cells were stored in sterile water at 4°C and GAD activity was intermittently tested over the course of one month.As shown in Fig.5,the GAD activity of the immobilized,permeabilized cells remained>90%during the 30-day testing period,which indicated that the immobilized,permeabilized cells had good stability.
4.Conclusions

Fig.3.Transmission electron micrographs of the recombinant E.coli.(a,c)Control(untreated cells);(b,d)cells treated with 5 μg·ml−1 ampicillin.
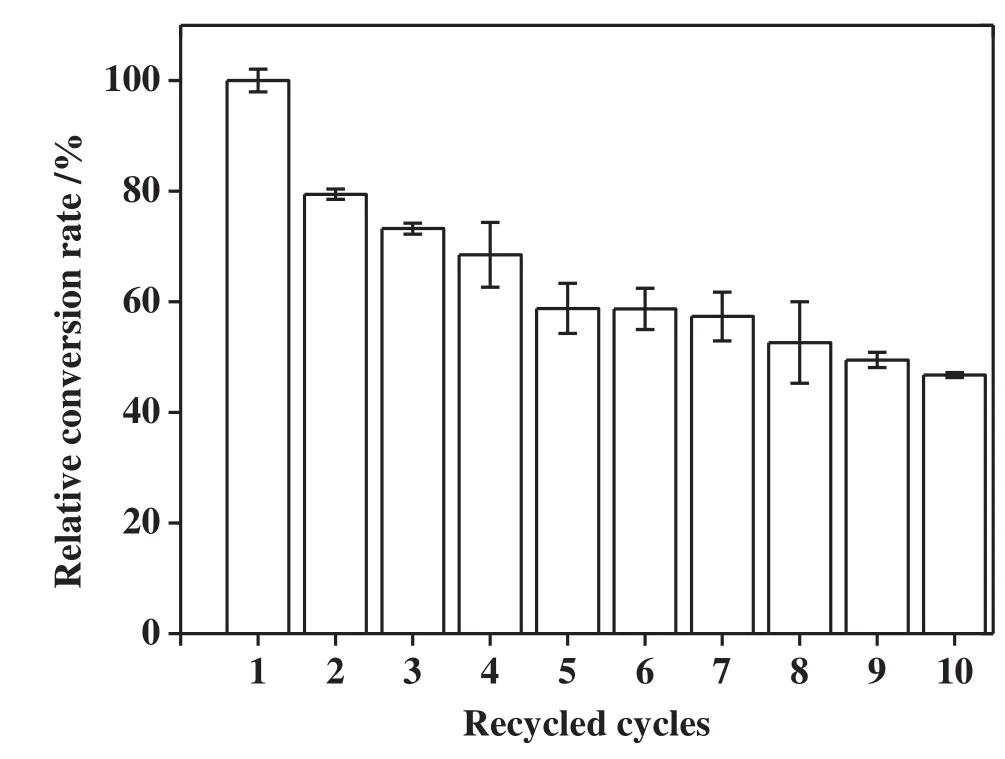
Fig.4.GABA production of 10 sequential cycles with immobilized permeabilized BL21(DE3)-pET28a-gadB cells at 37°C.Biotransformation of each cycle was performed in sodium acetate buffer(100 mmol·L−1,pH 4.4),100 mmol·L−1 L-MSG,0.1 mmol·L−1 PLP and 100 r·min−1 shaking for 8 h.
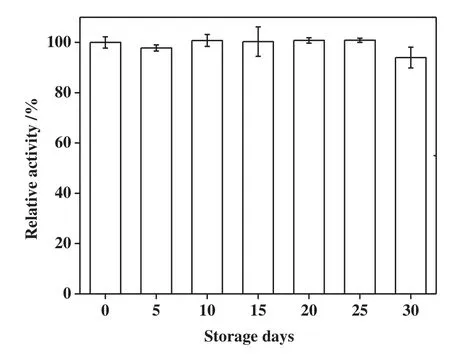
Fig.5.The storage stability of immobilized permeabilized BL21(DE3)-pET28a-gadB cells.The immobilized cells were stored at 4°C in sterile water and the GAD activity was intermittently tested over the course of one month.
Improving the cell-bound GAD activity of cells is an important challenge because of the expanding spectrum of GABA applications.In the present study,a permeabilization method not requiring extra steps after cell cultivation was developed to enhance the cell-bound GAD activity in E.coli.By culturing BL21(DE3)-pET28a-gadB cells in the presence of an appropriate amount of ampicillin,cell-bound GAD activity of the recombinant E.coli was enhanced.This improvement in cell-bound GAD activity was attributed to the increase in cell wall permeability.When the ampicillin-permeabilized cells were immobilized in PVA-SA-boric acid beads,the immobilized beads showed good reusability and storage stability.Based on these results,we conclude that the methods reported here may be utilized for industrial applications.
[1]K.Krnjevic,Chemical nature of synaptic transmission in vertebrates,Physiol.Rev.54(1974)418–540.
[2]K.Inoue,T.Shirai,H.Ochiai,M.Kasao,K.Hayakawa,M.Kimura,H.Sansawa,Bloodpressure-lowering effect of a novel fermented milk containing γ-aminobutyric acid(GABA)in mild hypertensives,Eur.J.Clin.Nutr.57(2003)490–495.
[3]C.Jakobs,J.Jaeken,K.M.Gibson,Inherited disorders of GABA-metabolism,J.Inherit.Metab.Dis.16(1993)704–715.
[4]T.Okada,T.Sugishita,T.Murakami,H.Murai,T.Saikusa,T.Horino,A.Onoda,O.Kajimoto,R.Takahashi,T.Takahashi,Effect of the defatted rice germ enriched with GABAfor sleeplessness,depression,autonomic disorder by oral administration,J.Jpn.Soc.Food Sci.47(2000)596–603.
[5]H.Hagiwara,T.Seki,T.Ariga,The effect of pre-germinated brown rice intake on blood glucose and PAI-1 levels in streptozotocin-induced diabetic rats,Biosci.Biotechnol.Biochem.68(2004)444–447.
[6]T.M.Lammens,M.Franssen,E.L.Scott,J.Sanders,Synthesis of biobased N-methylpyrrolidone by one-pot cyclization and methylation of gammaaminobutyric acid,Green Chem.12(2010)1430–1436.
[7]C.Takahashi,J.Shirakawa,T.Tsuchidate,N.Okai,K.Hatada,H.Nakayama,T.Tateno,C.Ogino,A.Kondo,Robust production of gamma-amino butyric acid using recombinant Corynebacterium glutamicum expressing glutamate decarboxylase from Escherichia coli,Enzym.Microb.Technol.51(2012)171–176.
[8]I.Saskiawan,Biosynthesis of polyamide 4,A biobased and biodegradable polymer,Microbiol.Indones.2(2008)119–123.
[9]Q.Wang,Y.Q.Xin,F.Zhang,Z.Y.Feng,J.Fu,L.Luo,Z.M.Yin,Enhanced gammaaminobutyric acid-forming activity of recombinant glutamate decarboxylase(gadA)from Escherichia coli,World J.Microbiol.Biotechnol.27(2011)693–700.
[10]J.Huang,L.H.Mei,H.Wu,D.Q.Lin,Biosynthesis of γ-aminobutyric acid(GABA)using immobilized whole cells of Lactobacillus brevis,World J.Microbiol.Biotechnol.23(2007)865–871.
[11]H.Ueno,Enzymatic and structural aspects on glutamate decarboxylase,J.Mol.Catal.B Enzym.10(2000)67–79.
[12]K.Park,S.Oh,Enhancement of γ-aminobutyric acid production in chungkukjang by applying a Bacillus subtilis strain expressing glutamate decarboxylase from Lactobacillus brevis,Biotechnol.Lett.28(2006)1459–1463.
[13]A.Y.Plokhov,M.M.Gusyatiner,T.A.Yampolskaya,V.E.Kaluzhsky,B.S.Sukhareva,A.A.Schulga,Preparation of gamma-aminobutyric acid using E.coli cells with high activity of glutamate decarboxylase,Appl.Biochem.Biotechnol.88(2000)257–265.
[14]D.L.V.Tam,T.W.Kim,S.H.Hong,Effects of glutamate decarboxylase and gammaaminobutyric acid(GABA)transporter on the bioconversion of GABA in engineered Escherichia coli,Bioprocess Biosyst.Eng.35(2012)645–650.
[15]L.Lin,S.Hu,K.Yu,J.Huang,S.J.Yao,Y.L.Lei,G.X.Hu,L.H.Mei,Enhancing the activity of glutamate decarboxylase from Lactobacillus brevis by directed evolution,Chin.J.Chem.Eng.22(2014)1322–1327.
[16]R.Chen,Permeability issues in whole-cell bioprocesses and cellular membrane engineering,Appl.Microbiol.Biotechnol.74(2007)730–738.
[17]Y.Numanoglu,S.Sungur,β-galactosidase from Kluyveromyces lactis cell disruption and enzyme immobilization using a cellulose-gelatin carrier system,Process Biochem.39(2004)703–709.
[18]A.Kumar,S.Singh,P.Poddar,A.Prabhune,A.Pundle,Effect of cultural conditions and media constituents on production of penicillin V acylase and CTAB treatment to enhance whole-cell enzyme activity of Rhodotorula aurantiaca(NCIM 3425),Appl.Biochem.Biotechnol.157(2009)463–472.
[19]P.Kaur,T.Satyanarayana,Improvement in cell-bound phytase activity of Pichia anomala by permeabilization and applicability of permeabilized cells in soymilk dephytinization,J.Appl.Microbiol.108(2010)2041–2049.
[20]A.De Leon,B.Garcia,A.de la Rosa,F.Villasenor,A.Estrada,R.Lopez-Revilla,Periplasmic penicillin G acylase activity in recombinant Escherichia coli cells permeabilized with organic solvents,Process Biochem.39(2003)301–305.
[21]M.Canovas,T.Torroglosa,J.L.Iborra,Permeabilization of Escherichia coli cells in the biotransformation of trimethylammonium compounds into L-carnitine,Enzym.Microb.Technol.37(2005)300–308.
[22]D.V.Cortez,I.C.Roberto,CTAB,Triton X-100 and freezing–thawing treatments of Candida guilliermondii:effects on permeability and accessibility of the glucose-6-phosphate dehydrogenase,xylose reductase and xylitol dehydrogenase enzymes,New Biotechnol.29(2012)192–198.
[23]B.Naji,G.Gehin,R.Bonaly,Structure of surfactants and glutamate ef flux by Corynebacterium glutamicum,Process Biochem.35(2000)759–764.
[24]A.Huchenq,M.Marquet,M.Welby,H.Montrozier,G.Goma,G.Laneelle,Glutamate excretion triggering mechanism—a reinvestigation of the surfactant-induced modification of cell lipids,Ann.Microbiol.(Paris)135B(1984)53–67.
[25]E.Y.Fan,J.Huang,S.Hu,L.H.Mei,K.Yu,Cloning,sequencing and expression of a glutamate decarboxylase gene from the GABA-producing strain Lactobacillus brevis CGMCC 1306,Ann.Microbiol.62(2012)689–698.
[26]Z.Q.Liu,M.Zhou,X.H.Zhang,J.M.Xu,Y.P.Xue,Y.G.Zheng,Biosynthesis of iminodiacetic acid from iminodiacetonitrile by immobilized recombinant Escherichia coli harboring nitrilase,J.Mol.Microbiol.Biotechnol.22(2012)35–47.
[27]R.Dave,D.Madamwar,Esterification in organic solvents by lipase immobilized in polymer of PVA-alginate-boric acid,Process Biochem.41(2006)951–955.
[28]S.Cheng,D.Wei,Q.Song,X.Zhao,Immobilization of permeabilized whole cell penicillin G acylase from Alcaligenes faecalis using pore matrix crosslinked with glutaraldehyde,Biotechnol.Lett.28(2006)1129–1133.
 Chinese Journal of Chemical Engineering2016年7期
Chinese Journal of Chemical Engineering2016年7期
- Chinese Journal of Chemical Engineering的其它文章
- Vanadium oxide nanotubes for selective catalytic reduction of NO x with NH3
- A unified graphical method for integration of hydrogen networks with purification reuse☆
- Optimal design for split-and-recombine-type flow distributors of microreactors based on blockage detection☆
- Theoreticalpredictions ofviscosity ofmethane under confined conditions☆
- Formation of crystalline particles from phase change emulsion:In fluence of different parameters
- The effect of SiO2 particle size on iron based F–T synthesis catalysts