
紫花苜蓿不同品种酯酶同工酶分析
2016-02-10 17:48赵菲佚焦成瑾康涛涛安建平天水师范学院生物工程与技术学院甘肃天水741001
天水师范学院学报 2016年2期
赵菲佚,焦成瑾,唐 红,康涛涛,安建平(天水师范学院 生物工程与技术学院,甘肃 天水 741001)
紫花苜蓿不同品种酯酶同工酶分析
赵菲佚,焦成瑾,唐红,康涛涛,安建平★
(天水师范学院 生物工程与技术学院,甘肃 天水 741001)
使用非变性聚丙烯酰胺凝胶电泳技术,对8个紫花苜蓿品种酯酶同工酶染色谱带、相似性指标进行了分析。结果表明,各品种酯酶同工酶在酶谱上均有表达,并呈现较强的特异性,8个品种存在一条共有同工酶条带。8个供试苜蓿品种共获得19条酶带,品种间酯酶同工酶表达存在显著差异。表达最多的品种有4条,最少为1条。部分苜蓿品种同工酶谱带存在相似性,而各同工酶的表达量存在差异。各品种酶谱联合系数计算结果表明:德宝和甘农三号的亲缘关系最远,新疆大叶和三得利之间的亲缘关系最近。
苜蓿;酯酶同工酶;电泳;酶谱联合系数;亲缘关系
紫花苜蓿为多年生豆科牧草,抗逆性强,适应范围广,能生长在多种类型的气候、土壤环境下,是世界上栽培最早、面积最大、经济价值最高的牧草,有“牧草之王”的美称,因此在我国被广泛种植。在生产实践中,由于在同一地区需种植不同品种的紫花苜蓿,或从其它地区进行引种进行种植示范,使得对于紫花苜蓿不同品种的鉴定成为必要。由于紫花苜蓿在形态外观上各不同品种差异不大,仅从形态很难对不同品种进行准确鉴别。近年来发展的分子标记鉴定方法,可从核酸水平上达到此目标。[1-3]然而,由于紫花苜蓿栽培种大多数在遗传上属于同源4倍体,且无法通过常规测序方法进行全基因组测序,目前主要使用RAPD法以筛选或建立组合分子标记方法进行不同品种的鉴定。此方法具有较好的鉴定效果,但也存在需要合成大量的随机引物,且需要昂贵的核酸电泳凝胶成像系统。因而在条件不具备的地区存在因难。酯酶同工酶是目前在品种鉴定中应用最多的酶类,酯酶(Ester⁃ase,EST)是具有多态性的一组同工酶,是生物体内广泛存在的水解酶类,酯酶同工酶是目前已发现的诸多同工酶中基因位点比较多(一般2~10个)的一种酶,在生物体内广泛存在,[4]已有研究表明,它在种系鉴定中具有重要的参考价值,[5]已广泛应用于鱼腥草[6]、菌株[7]、昆虫[8]等生物种的鉴定中。利用酯酶同工酶分析技术可以揭示群间或居群内甚至居群以下不同个体间或亲缘种之间的遗传上的变异,从而揭示同一个种内不同居群和同一个居群不同个体在遗传上的多样性。[9-10]因而以酯酶同功酶为标记,以聚丙烯凝胶电泳分析技术,通过对酯酶同工酶特征来鉴定不同的紫花苜蓿品种。为苜蓿的引种、育种、品种改良与栽培提供依据。
本研究使用同工酶电泳技术,对8个在我省进行种植的紫花苜蓿品种进行了多样性研究,目的在于研究8个供试紫花苜蓿品种间的多样性和亲缘关系。试验对8个紫花苜蓿品种的酯酶同工酶进行非变性凝胶电泳分析,考察不同品种的酯酶同工酶的表达状况,并对其进行聚类,为苜蓿的分型与育种提供依据。
1 材料与方法
1.1供试材料与培养
供试的8个紫花苜蓿品种阿尔冈金、润布勒、陇东苜蓿、三得利、甘农3号、德宝、新疆大叶和德福均来自天水农业科学研究所,其编号、中文名称、拉丁名称及原产地见表1.材料培养为将供试材料的种子取适量在50 ml烧杯中经6~8 h吸胀后种于人工培养土中,培养土的组成为:壤土:粗沙:泥炭土:蛭石=1:1:1:1,培养间温度为22~25℃,湿度50%~70%,16 h光,8 h暗培养。待长出3片真叶后取其叶片进行总蛋白提取。
1.2试验方法

表1 供试紫花苜蓿(Medicago sativa L.)材料及其来源
1.2.1叶片总蛋白提取
剪取8种供试苜蓿鲜叶各0.5 g置于预冷研钵中,加入少量石英砂,研磨过程中依次加入2 ml预冷0.1 M Tris-HCl缓冲液(pH 8.0),研磨样品匀浆后迅速移入预冷的2 ml离心管,4℃下12 000 rpm离心15 min,取8种不同品种苜蓿上清液0.8 ml置于新离心管中,4℃保存备用。
1.2.2总蛋白非变性聚丙烯酰胺凝胶电泳
试验使用非变性聚丙烯酰胺凝胶垂直板电泳法。[11]分离胶浓度为8%,浓缩胶为4%.电极缓冲液为Tris-甘氨酸电极缓冲液(pH 8.0)。提取总蛋白样品20 μl加入3 μl 6x蛋白载样缓冲液。初始电压60 V,约30 min后样品进入分离胶时,电压升至100 V,当溴酚蓝指示剂前沿移至分离胶底部约1.5 cm时停止电泳并剥胶。
1.2.3非变性聚丙烯酰胺凝胶染色
聚丙烯酰胺凝胶染色使用醋酸联苯胺法,将电泳后的凝胶浸入染色液(100 ml磷酸缓冲液染液中含50 mg乙酸-α-萘酯、50 mg乙酸-β-萘酯和200 mg坚牢蓝RR盐)中,室温下约20 S后出现染色酶带至清晰时停止染色过程,取出后用ddH2O漂洗,进行拍照。
1.2.4数据处理
酶谱带迁移率(Rf)计算:Rf=酶带迁移距离(cm)/溴酚兰迁移距离(cm)。[10]按酶带的相对迁移率、颜色深浅、宽度以及染色酶带数目标定酶带位置。[11]根据所统计酶谱带的有无和强弱,将酶谱条带分为强带、较强带、弱带和无条带。计算不同品种每种谱带的相对迁移率和相似系数。使用公式:S=2w/(a+b)×100%对8个苜蓿不同品种酯酶同工酶酶谱相似系数进行计算。公式中a,b,w分别代表a品种,b品种和a、b品种共有的酶带数,其中S以小数表示。
2 结果与分析
2.1紫花苜蓿不同品种EST同工酶分析
为考察不同紫花苜蓿品种的酯酶同工酶表达的差异,在前述试验的基础之上,对不同品种苜蓿的粗酶液进行了提取,经非变性聚丙烯酰胺凝胶电泳分离后,使用酯酶同工酶染色液进行染色,结果如图1所示。
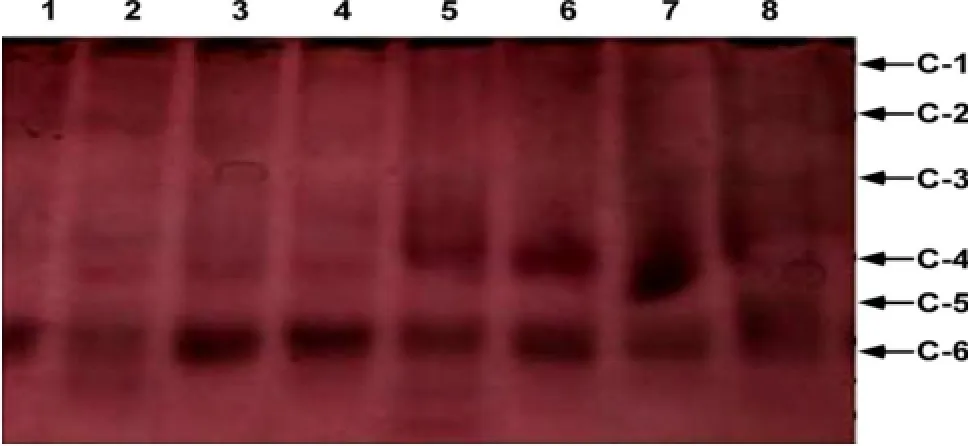
图1 8种供试苜蓿品种酯酶同功酶酶谱分布
图1结果表明,8种供试苜蓿品种共呈现19条酶带,表明8种苜蓿中酯酶同工酶均有表达。在6种不同迁移率的同工酶带中,C-4条带为8种苜蓿共有,品种3、4、5、6、7、8缺失C-1条带,品种1、2、3、4、8缺失C-2带,品种1、5、6、7、8无C-3带,C-5带为品种2、5共有,C-6带为品种5所独有。供试苜蓿品种共有一条主酶带,其中品种2、5各自出现4条酶带,品种8出现1条酶带,品种1、3、4、6、7各自出现2条酶带。供试苜蓿品种出现不同的酶带,且其同工酶的活性也存在差异,此表明8种苜蓿中的酯酶同工酶在遗传上存有差异。品种3酶带着色最深、条带最宽,预示其酯酶同工酶活性最强,其他7种苜蓿酯酶酶带着色较浅,酯酶同工酶活性较低,且品种间存在差异,呈现酯酶同工酶在不同的苜蓿品种间具有多态性。
2.2各供试材料酶带相对迁移率的计算
以苜蓿酯酶同工酶分析为基础,为考察不同品种间的亲缘关系,对所获得的酯酶同工酶谱带计算其相对迁率(Rf),结果如表2所示。
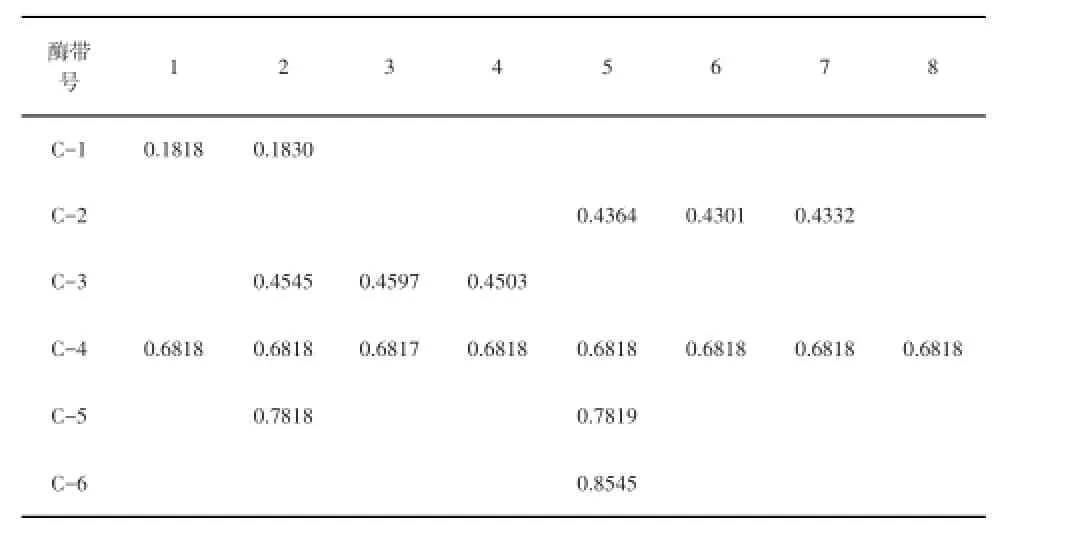
表2 酯酶同功酶酶带相对迁移率
由表2可知,8个紫花苜蓿品种酯酶同工酶谱带迁移率(Rf)在0.1818~0.8545之间,共检测到19条酶带,1条特征谱带;酶带条数最多的是品种5,有4条;酶带数量最少的是品种8,只有1条带。另外,在8种紫花苜蓿中,C-6仅为品种5所特有。各苜蓿不同品种酯酶谱带相互区别,可作为品种鉴定的特征酶带。
2.3酶谱联合系数S的计算
为考察苜蓿不同品种间的亲缘关系,对酯酶同工酶计算酶谱联合系数。计算结果如表3所示。
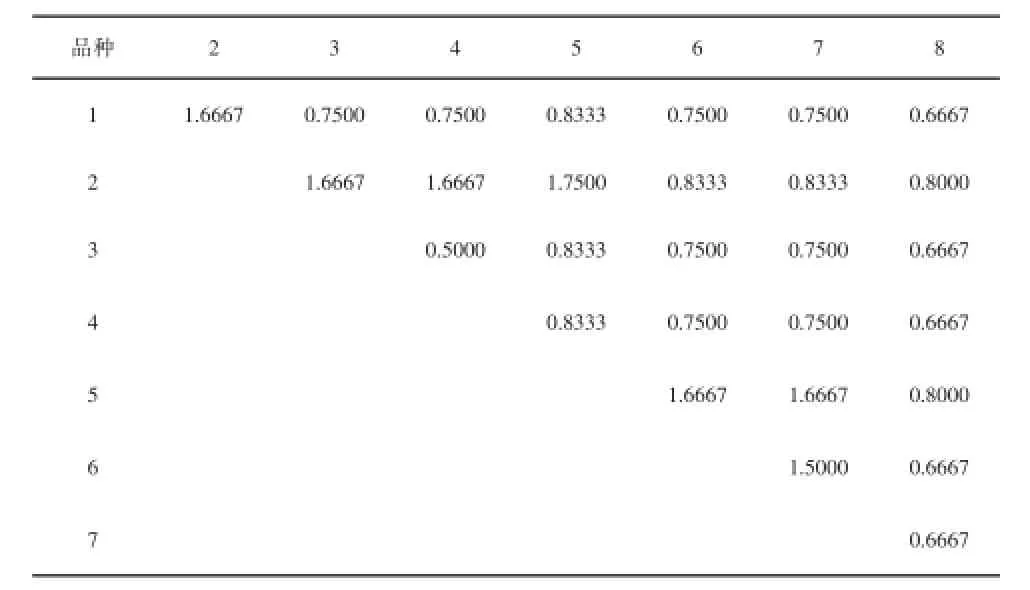
表3 苜蓿不同品种酯酶酶谱联合系数矩阵表
由表3可知,联合系数S在0~2之间变化,其值越大,两品种亲缘关系越近,越小亲缘关系越远。品种3和品种4的联合系数最小(S=0.5000),品种6和品种7之间的联合系数为1.5000,它们的酶带数均为2条,酶带位置也相同,但其酶活性存在差异。品种1和品种2之间,品种2和品种3、4之间,品种5和品种6、7号之间的联合系数均为1.6667,亲缘关系十分相近。其余几个品种之间亲缘关系介于上述亲缘关系之间。表明其亲缘介于上述品种之间。
3 讨论
本研究使用非变性PAGE对8个苜蓿品种酯酶同功酶进行了分析,结果可检测到共19条酯酶同工酶带,有6种迁移率不同的酶带。8种苜蓿中酯酶同工酶均可得到表达。此外,联合系数计算和分析表明,德宝和甘农三号的亲缘关系最远,新疆大叶和三得利之间的亲缘关系最近。
8个苜蓿品种酯酶同工酶在不同的苜蓿品种间,存在不同品种间的共有酶带,也同样具有单个品种的特有酶带,使得不同品种间的谱带可进行区别。同工酶酶谱具有较高的多态性水平,品种间相似系数越小则表明该品种间亲缘关系越远,品种3和品种1这两个品种间亲缘关系最远。8个苜蓿品种的Rf值介于0.1818~0.8545之间且都具有一条主条带,显示出其遗传稳定性,6条不同迁移率的酶带表现其多态性现象。8个品种的酯酶同工酶在酶带数、各条带染色强度上存在差异,表明品种之间存在遗传差异,经过长期演化,亲缘关系有所改变,各自在特定的生长环境或不同的进化过程中已发生趋异分化,[15]这可作为苜蓿种间鉴定的一个标志。
酯酶同工酶酶带数目、迁移率Rf及酶活性上的差异,可用于了解某一植物组织或器官的某些同工酶带在一定发育时期的出现以及在有外界环境条件影响时这些酶带减弱或消失,或者在不同品种植物中同工酶也表现出不同的特征,这种现象说明分化基因作用和环境条件影响都会使植物产生生理生化上的差异。[12-13]因而,在使用酯酶同工酶技术鉴定植物不同品种时也应通过形态学,细胞学等研究相结合而最终予以确认结果的正确性。发展更加简便、快速和准确的以同功酶为基础的品种鉴定方法是未来研究的重点。
[1]蒋素梅,冯莉,林德球.14个黄皮品种(系)的RAPD分析[J].亚热带植物科学,2008,37(2):26-29.
[2]黄海,李劲松,曹兵.分子标记技术在石斛属植物种质资源研究中的应用[J].生物技术通报,2010,(4):75-87.
[3]王学军,熊兴华,官春云.利用RAPD技术对5个油菜(B.napus)品种进行鉴别[J].作物研究,2010,24(1):99-102.
[4]袁带秀,侯娟.3个不同种源地三叶青酯酶同工酶比较研究[J].湖南农业科学,2010,(21):34-36.
[5]汤加勇,李青苗,杨瑞武,廖进秋,周永红.6种郁金类药用植物的同工酶研究[J].中国中药杂志,2008,33(12):1381-1386.
[6]吴卫,郑有良,陈黎,杨瑞武,颜泽洪,魏育明.川产鱼腥草种质资源的同工酶分析[J].栽培与育种,2002,25(10):695-698.
[7]王俊玲,李明,田景花,张成合,安振营.6个杏鲍菇菌株及其杂交子代的酯酶同工酶分析[J].河北农业大学学报,2004,27(3):29-32.
[8]郭晓霞,郑哲民,于广志.酯酶同工酶多态性及其在昆虫分类学中的应用价值[J].昆虫知识,2000,37(6):371-374.
[9]GOTTLIEB L D.Gene number in species of Astereae that have different chromosome number[J].Proc Natl Acad Sci.USA,1981,78(9):3726-3729.
[10]李景环,云锦凤,王树彦,等.酯酶同工酶标记坚定加拿大披碱草和老芒麦的杂种后代纯种研究[J].种子学报,2007,26(11):75-76.
[11]郭尧君.蛋白质电泳实验技术[M].北京:科学出版社,1999:128-131.
[12]郭宝生,翁跃进.大豆耐盐机理及相关基因分子标志[J].植物学通报,2004,21(1):113-120.
[13]刘波,王荔,陈疏影,等.36份不同居群半夏同工酶研究[J].云南农业大学学报,2008,23(1):11-14.
〔责任编辑 王小风〕
S551+.703
A
1671-1351(2016)02-0019-03
2016-01-23
赵菲佚(1972-),男,江苏镇江人,天水师范学院生物工程与技术学院副教授,博士。
安建平(1965-),男,甘肃秦安人,天水师范学院生物工程与技术学院教授。
国家自然科学基金(31260568,31160060)及天水市科技局2010年科技支撑计划项目阶段性成果
猜你喜欢
微生物学杂志(2020年6期)2020-03-05
天然产物研究与开发(2018年8期)2018-09-10
天然产物研究与开发(2018年4期)2018-05-07
分析化学(2017年12期)2017-12-25
江苏农业科学(2016年8期)2017-02-15
浙江大学学报(工学版)(2016年2期)2016-06-05
烟草科技(2015年8期)2015-12-20
河北渔业(2015年9期)2015-09-15
中国当代医药(2015年21期)2015-03-01
郑州大学学报(医学版)(2015年1期)2015-02-27
