
饥饿诱导下小鼠成骨细胞自噬与凋亡相互作用研究
2016-01-12 06:11
川北医学院学报 2015年2期
饥饿诱导下小鼠成骨细胞自噬与凋亡相互作用研究
朱文胤1,2,陈倩倩1,刘祁2,周云3,晏燕4,曹军1
(1.第四军医大学口腔医院正畸科,军事口腔医学国家重点实验室,陕西西安710032; 2.解放军75220部队医院,广东潮州
521000;3.解放军第461医院口腔正畸科,吉林长春130000;4.川北医学院附属医院,四川南充637000)
【摘要】目的:探讨饥饿条件下自噬与凋亡在小鼠成骨细胞中的发生情况与相互关系。方法:培养初代MC3T3-E1小鼠成骨细胞系,将其分为饥饿诱导组与3-甲基腺素(3-MA)预处理饥饿诱导组。其中,饥饿诱导组采用单纯饥饿诱导法,用厄尔氏平衡盐溶液(earle’s balanced salt solution,EBSS)培养1 h、2 h、3 h、4 h、5 h、6 h。3-MA是一种特异性自噬抑制剂,在3-MA预处理饥饿诱导组中,正常培养的小鼠成骨细胞加入3-MA预处理1 h后,再用EBSS平衡盐溶液培养1 h、2 h、3 h、4 h、5 h、6 h。应用透射电子显微镜观察饥饿诱导组小鼠成骨细胞自噬与凋亡的形态学变化,应用Western blot免疫印迹法和Annexin V-FITC/PI双标记流式细胞术检测小鼠成骨细胞中自噬相关蛋白LC3,凋亡相关蛋白caspase 3表达水平的变化。结果:透射电镜结果显示单纯饥饿诱导条件下,细胞在2 h前以自噬为主,2 h后以凋亡为主,并随着时间的延长,凋亡水平逐渐增高。Western blot免疫印迹法结果显示与饥饿诱导组相比,3-MA预处理饥饿诱导组的LC3转化水平在1 h、2 h、3 h、4 h、5 h、6 h都出产生了明显的下降趋势(P<0.05); caspase 3蛋白表达水平在1 h、2 h、3 h和4 h增高(P<0.05),但0 h、5 h和6 h无差异(P>0.05)。Annexin V-FITC/PI双标记流式细胞术检测结果和单纯饥饿诱导组相比,3-MA预处理组仅在1 h、2 h、3 h和4 h促进了细胞凋亡(P<0.05),而并未影响到0 h、5 h和6 h的细胞凋亡(P>0.05)。结论:饥饿发生后,小鼠成骨细胞的自噬作用通过降低caspase 3的活化水平,在细胞饥饿2 h前拮抗凋亡作用,随着饥饿时间的不断延长,细胞自噬的表达水平在3 h 和4 h时不断下降,自噬对细胞的保护不断降低,最终走向凋亡,此时自噬对凋亡的发生将不再有任何作用。
【关键词】成骨细胞;自噬;凋亡;饥饿
牙周组织改建是正畸牙齿移动的基础,有研究表明:在施加正畸矫治力初期,牙周组织发生改建,压力侧牙槽骨在正畸力作用下管径减小,毛细血管数及血供减少,从而使压力侧牙槽骨成骨细胞处于一个低氧、营养缺乏的饥饿环境中,这种刺激可诱导细胞发生死亡[1]。细胞的正常死亡包括两种方式:自噬性程序性死亡和凋亡性程序性死亡[2]。然而,自噬和凋亡在细胞死亡的过程中所扮演的角色目前还存在争议。有研究报道自噬通过分解代谢为细胞提供养分拮抗凋亡,延长细胞在代谢应激期间的存活;但同时也有研究证实自噬作为一种死亡机制协同凋亡促进细胞死亡,在不同的细胞中及不同的状态下自噬将发挥不同的作用[3-4]。在正畸压力侧牙槽骨成骨细胞的死亡中,自噬到底抑制了成骨细胞的生存还是促进了细胞死亡都尚不明确;另一方面,成骨细胞在饥饿死亡过程中,自噬、凋亡的发生情况及相互关系目前未见文献报道[5]。厄尔氏平衡盐溶液(Earle's balanced salt solution,EBSS)饥饿诱导法是目前自噬诱导的经典方法,EBSS平衡盐缓冲液内无氨基酸与血清,能维持细胞一定的代谢基本需要,是饥饿诱导自噬发生常用试剂[6]。3-甲基腺素(3-MA)是一种特异性自噬抑制剂[7],可通过与常规饥饿诱导细胞的对比,更好的表明自噬与凋亡的相互关系。本研究通过饥饿诱导条件下成骨细胞自噬与凋亡表达的水平变化和相互关系,初步分析自噬和凋亡对正畸牙齿移动过程中牙槽骨改建的作用,为完善正畸力作用下牙周组织改建的生物学机制提供参考。
1 材料与方法
1.1主要仪器与试剂
主要仪器: mini protean 3 cell电泳仪购自美国BIO-RAD公司,Olympus-BH-2型光学显微镜购自日本Olympus公司,流式细胞仪购自美国Becton Dickson公司,Zeiss Axioimager M1型显微镜购自德国ZEISS公司。主要试剂:新生胎牛血清、胰蛋白酶、α-MEM培养基购自HyClone公司; 3-甲基腺素(3-MA)、EBSS由Gibco公司购买;鼠抗人多克隆抗体微管相关蛋白l轻链3(LC3)、鼠抗人多克隆抗体caspase3(含半胱氨酸天冬氨酸蛋白水解酶)购自美国Cell Signaling Technology公司; Annexin V/PI试剂盒从南京凯基生物制品公司购买; RIPA细胞裂解液、山羊抗兔HRP-标记二抗自上海基尔顿公司购得。
1.2传代培养MC3T3-E1小鼠成骨细胞
小鼠胚胎成骨细胞MC3T3-E1初代由ATCC购买,α-MEM培养液中加入HyClone 10%胎牛血清作为细胞培养基,细胞培养箱各项指标: 37℃、CO2含量(为5%)、饱和湿度。当细胞在培养皿中生长于镜下观察处于对数生长期后,去除原培养液,预热PBS漂洗,以EBSS培养液诱导细胞饥饿状态。3-MA预处理组细胞饥饿前1 h加入5 mmol/L 3-MA预处理,用以抑制饥饿诱导的自噬。
1.3实验分组
取生长良好的小鼠成骨细胞随机分为两组: EBSS饥饿诱导组以EBSS培养液替代常规细胞培养液进行培养,饥饿诱导不同时间,分别为1 h、2 h、3 h、4 h、5 h、6 h。3-MA预处理组细胞饥饿诱导前1 h加入5 mmol/L 3-MA预处理,随后用EBSS培养液培养,分别进行饥饿诱导1 h、2 h、3 h、4 h、5 h、6 h。两组组内空白对照均为含10%胎牛血清的α-MEM培养基培养正常小鼠成骨细胞。
1.4透射电子显微镜观察饥饿诱导组小鼠成骨细胞自噬体、凋亡小体的出现顺序和数量的变化
取EBSS饥饿诱导各组细胞,胰蛋白酶消化离心,加入戊二醛固定12 h以上,锇酸再次固定,醋酸铀染色,梯度乙醇脱水,树脂包埋,用LKB-V超薄切片机以0.1 um厚度切片,最后经柠檬酸铅染色,于透射电子显微镜下观察细胞各细胞器结构,分析各分组中自噬体和凋亡的典型形态学变化。
1.5Annexin V/PI双标流式细胞术检测细胞凋亡
各组实验处理后的细胞离心、收集、漂洗,加入Binding Buffer制备成细胞悬液。加入5 μL Annexin V-FITC混匀后再加入10 μL PI,室内温度下避光反应5~15 min。最后再加入Binding Buffer,上机检测细胞凋亡状况。用流式细胞仪检测的激发波长Ex为488 nm,发射波长Em为530 nm。经双变量流式细胞仪检测,其散点图结果中B1区为坏死细胞,在本实验中代表因自噬而导致死亡的细胞; B2区表示凋亡晚期细胞; B3区表示正常活细胞; B4区表示凋亡早期细胞。每组实验重复3次。凋亡细胞所占百分比可用以下公式计算获得:凋亡细胞所占百分率= (B2 + B4)%。
1.6Western blot检测LC3-Ⅱ/LC3-Ⅰ、caspase3表达水平变化
取各组实验处理后细胞,加入适量蛋白酶抑制剂(PMSF),然后重悬于细胞裂解液(RIPA)中,冰上裂解20 min同时超声裂解3次,以12 000 rpm、4℃条件下离心15 min,吸取的上清液为总蛋白液,将其移入1.5 mL离心管中,蛋白总浓度用BCA法测定,SDS-PAGE电泳分离;然后转移至聚偏氟乙烯(PVDF)膜上,加入BSA稀释后的一抗孵育过夜,5%脱脂奶粉洗去多余抗体,加入HRP标记的山羊抗兔IgG室温下摇振孵育2 h;最后加入ECL试剂,暗盒X光胶片显影,胶片经计算机扫描检测蛋白表达情况。所有条带灰度值输入Graph Pad软件进行分析,并形成图表。每组实验重复3次,分析不同分组间细胞自噬与凋亡表达水平的变化及其相互关系。
1.7统计学分析
实验所有数据输入SPSS19.0统计学软件分析,EBSS饥饿诱导组各时间点试验样本数据比较采用方差检验统计分析; EBSS饥饿诱导组与3-MA预处理组在同一时间点两组组间数据比较采用t检验统计分析; P<0.05认为差异有统计学意义。
2 结果
2.1透射电子显微镜观察小鼠成骨细胞超微结构结果
空白对照组小鼠成骨细胞胞核形态完整,核内明显核仁结构,细胞核位于细胞内一侧,胞浆内可见大量粗面内质网和高尔基复合体; 1 h组细胞胞浆内分布大量双层膜结构囊泡,囊泡内包裹线粒体、核糖体等细胞器,根据其形态结构可判定为为自噬体空泡;2 h组可见大量细胞出现凋亡染色质凝集,细胞核呈新月形边棘; 3 h组细胞内观察到凋亡凝集染色质和自噬小体共存; 4 h组可见大量凋亡细胞核; 5 h组凋亡细胞核破碎,出现凋亡小体;6 h组出现大量凋亡小体。由此可见,在EBSS饥饿诱导条件下,小鼠成骨细胞死亡方式在2 h前以自噬为主,2 h后以凋亡为主,并随着时间的延长,凋亡水平逐渐增高(图1)。
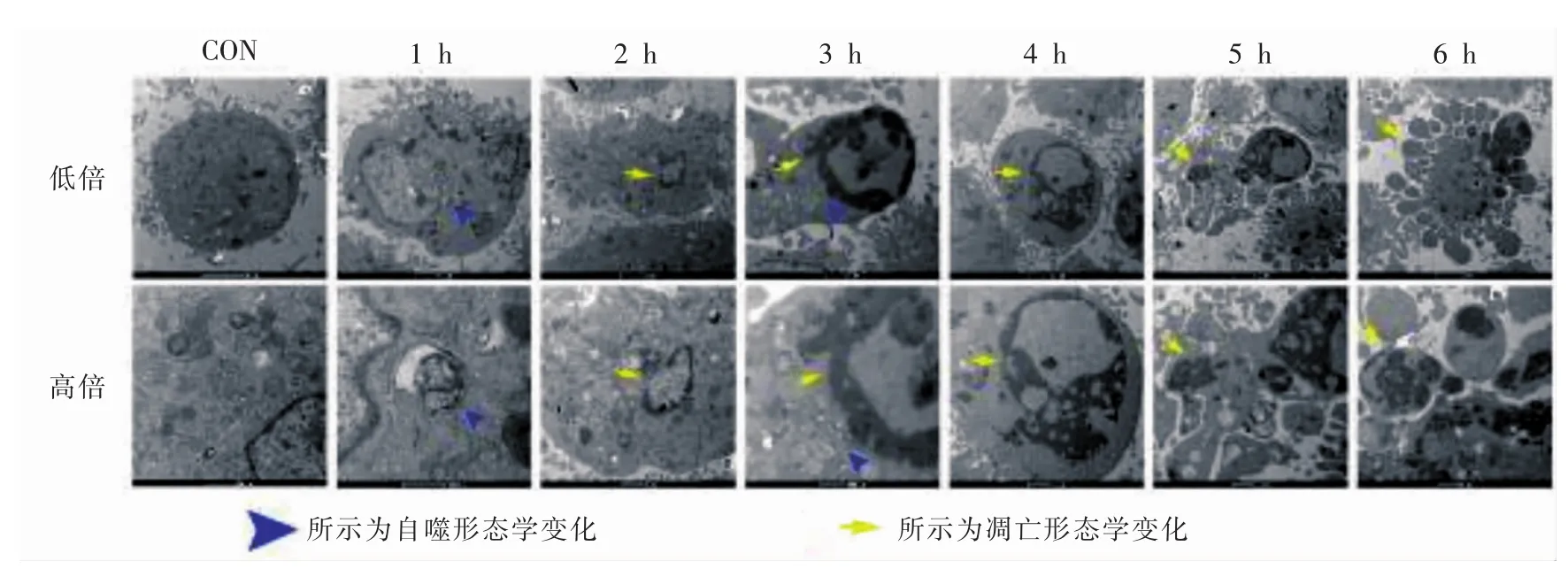
图1 小鼠成骨细胞的超微结构图
2.2Western blot免疫印迹检测结果
Western blot实验显示,饥饿会同时导致细胞产生自噬和凋亡现象,两者之间存在着以时间为联系的相互关系。其中各EBSS饥饿诱导组LC3-Ⅱ/LC3-Ⅰ比值均大于空白对照组(P﹤0.05),经EBSS饥饿诱导1 h后LC3-Ⅱ/LC3-Ⅰ比值开始上升,2 h组LC3-Ⅱ/LC3-Ⅰ比值到达最大值(P﹤0.05),自噬达到高峰,随后自噬水平开始下降(P﹤0.05),caspase3蛋白表达水平随时间逐渐升高(P﹤0.05),明显的凋亡高峰在4 h当自噬下降时被观察到(图2)。与EBSS饥饿诱导组相比,3-MA预处理组的LC3-Ⅰ向LC3-Ⅱ转化水平在除0 h外的其他时间点均出现明显下降(P<0.05)。比较caspase3蛋白表达水平发现,3-MA预处理组与单纯饥饿诱导组相比仅上调了1 h、2 h、3 h和4 h的caspase3蛋白表达水平(P<0.05),而并不影响0 h、5 h和6 h的caspase3蛋白表达水平(P>0.05),见图3。

图2 单纯饥饿诱导不同时间小鼠成骨细胞自噬与凋亡变化水平Western blot检测结果与分析

图3 3-MA预处理组与EBSS饥饿诱导组自噬与凋亡变化水平Western blot检测结果分析与对比
2.3Annexin V-FITC/PI双标记流式细胞术检测结果
空白对照组细胞凋亡量均小于各饥饿诱导组(P﹤0.05),各饥饿诱导组的细胞凋亡量随时间的上升而增高,6 h达到最大值(P﹤0.05)(图4)。比较3-MA预处理组和EBSS饥饿诱导组的细胞凋亡量(B2 + B4)%发现,3-MA预处理仅在1 h、2 h、3 h和4 h促进了细胞凋亡(P<0.05),而并未影响到0 h、5 h和6 h的细胞凋亡(P>0.5),这与之前的Western blot检测结果一致(图5)。
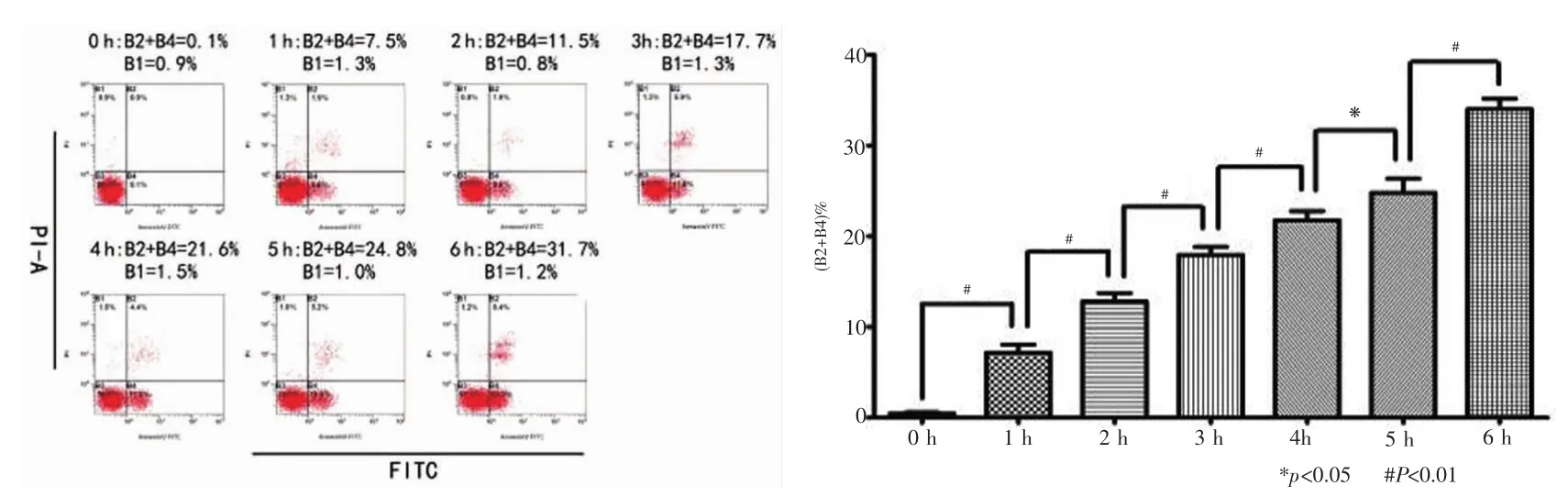
图4 EBSS饥饿诱导组Annexin V-FITC/PI双标法检测凋亡结果与分析
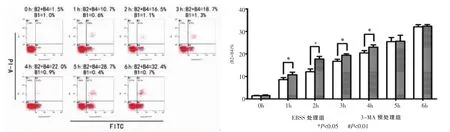
图5 3-MA预处理组与EBSS饥饿诱导组细胞Annexin V-FITC/PI双标法检测凋亡结果分析与对比
3 讨论
细胞死亡包括3种类型:自噬、凋亡和坏死[2]。凋亡属于I型程序性细胞死亡,依赖于caspase,表现为细胞间连接消失,细胞体积缩小,胞质密度增加,线粒体通透性改变,核质浓缩,核膜核仁破碎,DNA降解,最终分隔为几个凋亡小体被巨噬细胞所吞噬[2]。自噬性细胞死亡属于II型程序性细胞死亡,以自噬体的出现为特征,不依赖于caspase,自噬是指细胞内的溶酶体降解自身细胞器和其他大分子的过程[8-9]。自噬对细胞内环境的稳态和细胞生存至关重要,当细胞在缺乏营养或发生应激反应时,可诱发自噬现象[10-11]。自噬和凋亡无论在形态学还是生化代谢途径方面都有着明显的区别,但有研究证实,二者在功能上存在联系,即自噬为凋亡所需,自噬通常发生在凋亡之前,进而启动凋亡;自噬也可抑制凋亡,保护细胞免于发生凋亡和坏死;自噬可以向凋亡转化,共同促进细胞死亡[3-4]。目前研究显示,自噬与肿瘤、感染性疾病、自身免疫性疾病等许多病变有密不可分的关系[12-14]。Foster[15]认为矫治力作用后,压力侧牙周膜内液压升高,牙周膜血管内的血液减少,达到液压平衡,最后压力侧血管狭窄,血流量减少,同时该区域血管密度和面积均减小。那么当施加正畸矫治力导致压力侧血供减少,营养缺乏,成骨细胞处于饥饿环境,继而发生死亡。成骨细胞的死亡过程中自噬与凋亡的发生情况如何?两者是怎样的关系?针对这些问题本研究通过饥饿诱导法,使MC3T3-E1细胞处于饥饿环境,诱导自噬与凋亡的发生,并使用3-MA自噬抑制剂抑制自噬,观察自噬对凋亡的影响,利用透射电子显微镜定性分析小鼠成骨细胞自噬与凋亡的形态学变化,应用Western blot免疫印迹法和Annexin V-FITC/PI双标记流式细胞术定量分析小鼠成骨细胞中自噬相关蛋白LC3,凋亡相关蛋白caspase 3表达水平的变化。
从透射电子显微镜这一自噬检测金标准中观察到EBSS饥饿诱导组1 h可见自噬体,2 h出现凋亡染色质凝集,3 h观察到自噬体与凋亡细胞核共存,4 h、5 h、6 h细胞只出现凋亡小体,并随时间延长而增多。这证明了饥饿导致小鼠成骨细胞发生自噬与凋亡,早期以自噬为主,随着时间的延长逐渐走向凋亡。LC3蛋白作为自噬检测的的标志蛋白,LC3-I和LC3-II是它的两种不同表达形式,当细胞因外界刺激从而发生自噬时,细胞内LC3的总含量及LC3-I 向LC3-II的转化量将会明显增多; caspase3蛋白是真核细胞发生凋亡的标志性蛋白之一,与细胞的生长、分化及凋亡调节密切相关,当细胞发生凋亡,caspase3蛋白含量增高。Western blot实验结果显示饥饿同时诱导细胞产生自噬和凋亡反应,两者之间存在着以时间为联系的相互关系。在饥饿2 h期间,自噬达到高峰,随后自噬水平开始下降,明显的凋亡高峰在4 h当自噬下降时被观察到。由TEM和WB结果,我们已经知道饥饿引起的细胞死亡呈时间依赖性升高,为了进一步弄清自噬和凋亡在细胞死亡中所起的作用,明确在饥饿引起的细胞死亡中,自噬有没有起到作用,引入了Annexin V FITC/PI双标法,通过分析细胞坏死率,发现在饥饿诱导各分组都没有发现细胞坏死性死亡率有明显变化(P>0.05),因为自噬小体的产生是自噬性细胞死亡的典型形态特征。综合以上数据我们可以得知饥饿导致的细胞死亡中,自噬性细胞死亡并不占主导地位。因为自噬与凋亡在发生时间上具有先后顺序性,这一顺序性说明饥饿早期自噬对凋亡的发生可能具有拮抗作用。为了验证这一假设,引入了3-MA预处理细胞抑制自噬,重新检测LC3和caspase3活化水平,Western blot实验结果显示3-MA预处理组的LC3-Ⅱ/LC3-Ⅰ比值与单纯饥饿诱导组相比,在除0 h外的其他时间点下降趋势明显,比较caspase3蛋白表达水平发现3-MA预处理组与单纯饥饿诱导组相比仅上调了1 h、2 h、3 h和4 h的caspase3蛋白表达水平,而并不影响0 h、5 h和6 h的caspase3蛋白表达水平。Annexin VFITC/PI双标记流式细胞术检测结果显示比较3-MA预处理组和单纯饥饿诱导组的细胞凋亡量(B2 + B4)%发现,3-MA预处理仅在1 h、2 h、3 h和4 h促进了细胞凋亡,而并未影响到0 h、5 h和6 h的细胞凋亡,这与之前的Western blot检测结果一致。本实验结果证明当饥饿发生后,小鼠成骨细胞首先启动自噬机制,在2 h前拮抗caspase 3的活化,避免凋亡发生,随着饥饿时间的延长,饥饿状态不断加剧,细胞自噬作用在3 h和4 h连续下降,自噬对细胞的保护作用也随之降低,细胞不可避免的发生凋亡,自噬对凋亡的影响最终消失。caspase3导致的凋亡一般被定义为由于内质网(endoplasmic reticulum,ER)压力不断增大从而诱导凋亡产生。细胞内的蛋白质合成和折叠都是在内质网内完成,而应激状况导致内质网蛋白折叠压力不断增大,非折叠蛋白增多。有研究证明自噬分解非折叠蛋白除了提供ATP外,还能缓解ER压力[16-17]。综合上述观点,我们推测在MC3T3-E1细胞中,在饥饿导致的应激状态下,非折叠蛋白增多,细胞通过自噬反应不断分解,从而延缓了凋亡的发生,可这种情况无法持久,细胞内部对非折叠蛋白的自噬作用一但达到饱和,凋亡即不可避免的随之来临。而细胞内部如何吞噬非折叠蛋白,又是通过何种机制来延缓凋亡发生的都还需要进一步研究。
参考文献
[1]Vandevska-Radunovic V,Kristiansen AB,Heyeraas KJ,et al.Changes in blood circulation in teeth and supporting tissues incident to experimental tooth movement[J].Eur J Orthod,1994,16(5):361-369.
[2]Schweichel JU,Merker HJ.The morphology of various types of cell death in prenatal tissues[J].Teratology,1973,7(3):253-266.
[3]Kang C,You YJ,Avery L.Dual roles of autophagy in the survival of Caenorhabditis elegans during starvation[J].Genes Dev,2007,21 (17):2161-2171.
[4]Espert L,Denizot M,Grimaldi M,et al.Autophagy is involved in T cell death after binding of HIV-1 envelope proteins to CXCR4[J].J Clin Invest,2006,116(8):2161-2172.
[5]周云,付秀莉,高远,等.饥饿诱导环境下人牙周膜成纤维细胞的自噬作用研究[J].陕西医学杂志,2014,43(2):152-154.
[6]Kroemer G,Marino G,Levine B.Autophagy and the integrated stress response[J].Mol Cell,2010,40(2):280-293.
[7]喻琴琴,杨俊,李馨欣.自噬及其抑制剂的研究进展[J].实用医学杂志,2013,29(17):2908-2910.
[8]Tsukada M,Ohsumi Y.Isolation and characterization of autophagydefective mutants of Saccharomyces cerevisiae[J].FEBS Lett,1993,333(1-2):169-174.
[9]Levine B,Kroemer G.Autophagy in the pathogenesis of disease [J].Cell,2008,132(1):27-42.
[10]Colell A,Ricci JE,Tait S,et al.GAPDH and autophagy preserve survival after apoptotic cytochrome c release in the absence of caspase activation[J].Cell,2007,129(5):983-997.
[11]Lee J,Giordano S,Zhang J.Autophagy,mitochondria and oxidative stress: cross-talk and redox signalling[J].Biochem J,2012,441 (2):523-540.
[12]Friedhuber AM,Chandolu V,Manchun S,et al.Nucleotropic doxorubicin nanoparticles decrease cancer cell viability,destroy mitochondria,induce autophagy and enhance tumour necrosis[J].J Pharm Pharmacol,2015,67(1):68-77.
[13]Marcuzzi A,Bianco AM,Girardelli M,et al.Genetic and functional profiling of Crohn's disease: autophagy mechanism and susceptibility to infectious diseases[J].Biomed Res Int,2013,doi: 10.1155/2013/297501.
[14]Rovetta AI,Pena D,Hernandez DPR,et al.IFNG-mediated immune responses enhance autophagy against Mycobacterium tuberculosis antigens in patients with active tuberculosis[J].Autophagy,2014,doi:10.4161/15548627.2014.981791.
[15]Foster TD.A textbook of orthodontics[M].Oxford: Blackwell,1975:356.
[16]Kim I,Xu W,Reed JC.Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities[J].Nat Rev Drug Discov,2008,7(12):1013-1030.
[17]Rasheva VI,Domingos PM.Cellular responses to endoplasmic reticulum stress and apoptosis[J].Apoptosis,2009,14 (8): 996-1007.
(学术编辑:刘英)
Study on the interaction between autophagy and apoptosis of MC3T3-E1 at starvation state
ZHU Wen-yin1,2,CHEN Qian-qian1,LIU Qi2,ZHOU Yun3,YAN Yan4,CAO Jun1
(1.State Key Laboratory of Military Stomatology,Department of Orthodontics,School of Stomatology,The Fourth Military Medical University,Xi’an,710032 Shaanxi;2.75220th Hospital of PLA,Chaozhou 521000,Guangdong;3.Department of Orthodontics,461th Department of Orthodontics,Changchun 130000,Jilin;4.Affiliated Hospital of North Sichuan Medical College,Nanchong 637000,Sichuan,China)
【Abstract】Objective: To explore the interaction between autophagy and apoptosis of MC3T3-E1 at starvation state.Methods: MC3T3-E1 of 3-MA group was pretreated with 3-MA for 1 hour before being cultured in earle’s balanced salt solution(EBSS)for another 6 hours,and the samples without 3-MA pretreatment were taken as starvation group.MC3T3-E1 of different groups were sampled hourly.The morphological change of MC3T3-E1 form starvation group was observed by TEM.To determine the expression of LC3 and caspase 3,western blot and flow cytometry were performed.Results: The observation of TEM showed that MC3T3-E1 from starvation group was dominated by autophagy in the first two hours of the 6-hours period cultured in EBSS,whereas apoptosis was the main process after the first two hours and the level increased with time.The observation of western blot showed that,compared with those from starvation group,MC3T3-E1 from 3-MA group presented a reduced LC3 expression level (P<0.05)throughout the 6-hours period,and anbook=207,ebook=81enhanced caspase 3 expression level in the first 4 hours (P<0.05),while there was no difference observed between the two groups (P>0.05)in the last 2 hours(P>0.05).The observation of flow cytometry showed that,compared with those from starvation group,MC3T3-E1 from 3-MA group presented an enhanced apoptosis level in the first 4 hours (P<0.05),while there was no difference observed between the two groups (P>0.05)in the last 2 hours(P>0.05).Conclusion: Under starvation condition,autophagy of MC3T3-E1 may antagonize apoptosis in the first 2 hours by inhibiting the activity of caspase 3.While the autophagy level reduces in the middle 2 hours,apoptosis may not be affected by autophagy.
【Key words】Osteoblast; Autophagy; Apoptosis; Starvation
通讯作者:曹军,E-mail: wykun@fmmu.edu.cn
作者简介:朱文胤(1983-),男,江苏南京人,硕士研究生,主要从事正畸学方面的研究。
收稿日期:2015-01-25
doi:10.3969/j.issn.1005-3697.2015.02.19
【文章编号】1005-3697(2015)02-0206-06
【中图分类号】R734.2
【文献标志码】A
网络出版时间: 2015-5-1 01∶33网络出版地址: http://www.cnki.net/kcms/detail/51.1254.R.20150501.1333.016.html
猜你喜欢
金桥(2021年10期)2021-11-05
小学生作文(低年级适用)(2019年3期)2019-04-04
中国现代医生(2016年27期)2016-12-21
糖尿病新世界(2016年16期)2016-12-09
科技视界(2016年11期)2016-05-23
中国实用医药(2016年7期)2016-03-17
文史春秋(2016年8期)2016-02-28
科技视界(2016年5期)2016-02-22
山东体育学院学报(2015年2期)2015-05-27
小说月刊(2014年8期)2014-04-19
