
昆虫资源在水产饲料中的应用研究进展
2016-01-10 02:56李森林徐歆歆
饲料工业 2016年22期
■吉 红 李森林 徐歆歆
(西北农林科技大学动物科技学院,陕西杨凌 712100)
随着集约化程度的日益加强,水产业对优质蛋白源的需求越来越大,而传统蛋白源(如鱼粉)存在资源短缺问题,其矛盾已显得愈发突出。因此,近年来国内外持续聚焦可替代鱼粉蛋白源的研究,以保障水产业的可持续发展(Barroso等,2014)。鉴于资源量及易得性等原因,用于替代鱼粉的大部分蛋白质来源于豆科植物、油料种子和谷物蛋白等非动物蛋白源,但是植物蛋白源存在诸多限制性因素,如适口性差、氨基酸不平衡、存在抗营养因子、纤维含量高等问题。
当前,昆虫作为人类和养殖动物潜在的蛋白源越来越受到关注。由于很多鱼类在自然环境下捕食昆虫,因此,将昆虫资源饲料化有着天然的基础。昆虫易于繁殖,生长迅速,饲料转化效率高,且可以利用生物废弃物进行养殖(Makkar等,2014)。昆虫养殖可以是部分(从卵到幼虫、蛹或成虫)过程,也可以是一个完整(从卵到卵)的过程。在日本冲绳,人们用工业化仓库养殖瓜实蝇,每周可以生产四千万幼虫(Mitsuhashi,2010)。目前可以养殖物种包括蝴蝶和飞蛾(鳞翅目,物种超过300个)、甲虫(鞘翅目,物种超过200个)、苍蝇和蚊子(双翅目,物种接近200个)、臭虫(异翅目,物种少于100个)、蜜蜂和黄蜂(膜翅目,物种少于100个)、蟋蟀和蝗虫(直翅目)、草蛉(脉翅目)、蟑螂(蜚蠊目)、白蚁(等翅目)和跳蚤(隐翅目)也有养殖(大约10~20个物种)。
本文综述了昆虫营养成分、利用方式及存在的问题和解决策略,涉及的种类包括蚕蛹、蝇蛆或蝇蛹、黄粉虫、双翅目昆虫黑水虻的幼虫或蛹、直翅目昆虫蝗科(蝗虫和蚱蜢)、蟋蟀科(蟋蟀)等,以期为水产饲料原料开发提供思路,提升饲料行业对昆虫资源的重视程度,为昆虫资源在水产饲料中的应用提供参考。
1 昆虫营养组成
1.1 蛋白质和氨基酸
昆虫体内含有较为丰富的蛋白质,一般粗蛋白含量为42%~63%,与豆粕相近,稍低于鱼粉(见表1)。如蝇蛆各生长阶段粗蛋白含量高达50%~65%(陈明等,2015),黄粉虫幼虫、蛹、成虫各阶段粗蛋白含量分别占干物质的47.7%~54.25%、55.23%~58.70%、63.19%~64.29%,显著高于豆粕、肉骨粉等原料(周元军,2005),柞蚕中的粗蛋白含量为67%~70%左右,含较多球蛋白和清蛋白,营养价值高,较其他蛋白源更易于消化吸收。有些昆虫粗蛋白含量高达干物质的80%,如黄蜂为81%等(田华等,2011)。许多昆虫能产生抗菌蛋白和干扰素,具有抗菌、抗病毒、提高免疫力等作用,有些甚至还能抑制肿瘤发展。
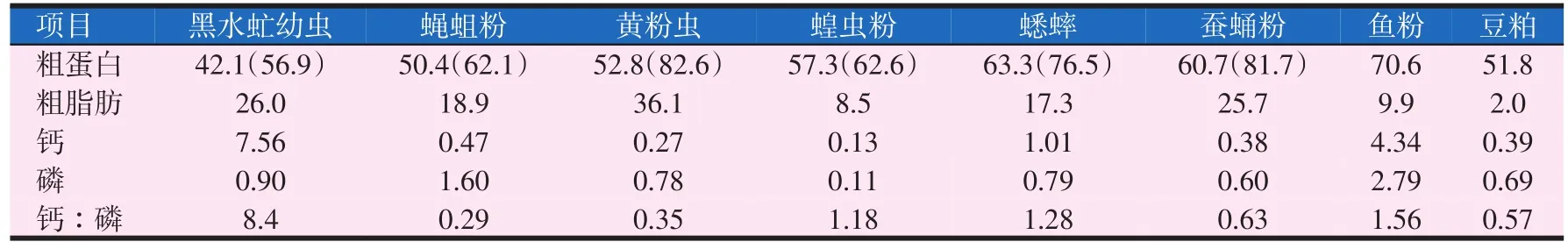
表1 昆虫粉、鱼粉和豆粕的主要化学组成(干物质,%)(Makkar等,2014)
对近千种昆虫体内氨基酸的分析表明,昆虫含有10%~30%的人体必需氨基酸(EAA),占氨基酸总含量的30%~50%。昆虫氨基酸种类和含量及其比例满足其做为饲料蛋白源所需的氨基酸模式(Schabel等,2010)。如表2所示,直翅目和黄粉虫赖氨酸含量低于鱼粉,而双翅目和家蚕赖氨酸较高,与鱼粉接近,高于豆粕。昆虫蛋氨酸含量通常低于鱼粉,苏氨酸和鱼粉相当,而家蚕的蛋氨酸、苏氨酸的含量较高,蝇蛆粉的色氨酸含量也比较高。胡金伟等(2009)发现烘干蝇蛆粉的蛋氨酸、亮氨酸、异亮氨酸含量高于鱼粉,其它种类氨基酸含量与鱼粉较为接近。黄粉虫蛹中必需氨基酸含量占总氨基酸的44.40%,含7种人体必需氨基酸。刘军等(2016)指出家蚕蛹、蛾的氨基酸组成基本一致,检出的氨基酸总量占蛋白质重的90%以上,其中必需氨基酸的总量约占氨基酸总量的38%。水产经济动物大都对昆虫的氨基酸有较高的消化利用率,最高可达98.93%,最低可达70%以上,接近或超过对畜禽肉、鱼粉的消化率,高于植物性蛋白的消化利用率。蚕蛹和双翅目氨基酸组成优于豆粕,更适合替代鱼粉。
1.2 脂肪和脂肪酸
昆虫脂肪含量较高,且包含水产动物所需的必需脂肪酸,其中不饱和脂肪酸含量比例较大。Ramos等(1997)分析了墨西哥78种食用昆虫的营养成分,发现其脂肪含量为4.2%~77.2%。吴时敏等(2001)发现昆虫在蛹、幼体和越冬期脂肪含量较成虫含量高。昆虫种类不同,其脂肪酸组成也不尽相同(见表3)。黄粉虫、蟋蟀和蝇蛆粉中不饱和脂肪酸含量较高(60%~70%),黑水虻幼虫中含量较低(19%~37%)。这可能是由于黑水虻油中月桂酸含量较高所致(21.4%~49.3%),而其它昆虫油脂不含或仅含少量月桂酸。和植物油(大豆油和葵花籽油)相似,通常昆虫的亚油酸(18∶2n-6)含量高于α-亚麻酸(APA,18∶3n-3)。昆虫油脂的脂肪酸组成受饲料、发育、种类等条件影响,一般含较多的棕榈酸(16∶0),硬脂酸(18∶0)的含量相对较低,单不饱和脂肪酸含量在20%~40%,以油酸(18∶1n-9)为主。多不饱和脂肪酸中亚油酸(18∶2n-6)和α-亚麻酸(18∶3n-3)较为突出。黄粉虫幼虫干粉的脂肪酸含量为66.28%,不饱和脂肪酸为77.05%,包括41.70%的亚油酸(武宝生等,2015)。中华稻蝗含较多的不饱和脂肪酸,并以油酸、亚油酸及亚麻酸为主要成分,约占脂肪酸总量的83%,油脂质量高于菜籽油和花生油(赵云涛等,2002)。刘军等(2016)经计算机质谱库检索,鉴定出雌雄蚕蛹油中8种脂肪酸,其中α-亚麻酸、油酸和棕榈酸3种脂肪酸的含量最高,其相对含量总和占到总脂肪酸含量的85%以上,而不饱和脂肪酸的相对含量均在70%左右,其中以α-亚麻酸最为突出。与鱼油相比,陆生昆虫n-6多不饱和脂肪酸含量较高,EPA(20∶5n-3)和DHA(22∶6n-3)含量较少。海水鱼需要较多的EPA和DHA,而自身合成能力有限,因此EPA和DHA是陆生昆虫应用于海水鱼饲料的限制性因素。
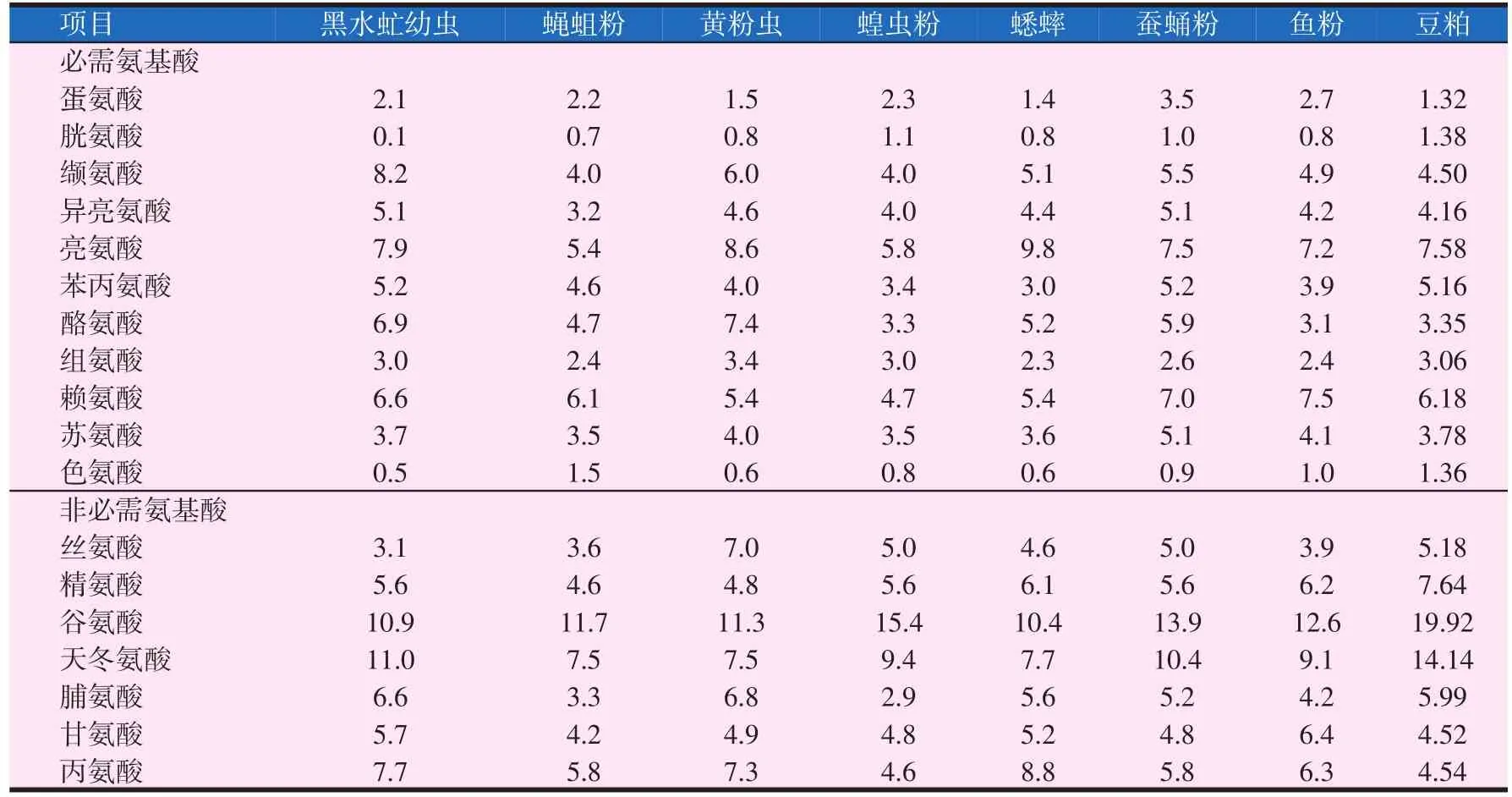
表2 昆虫粉、鱼粉和豆粕氨基酸组成(Makkar等,2014)(g/16 g氮)
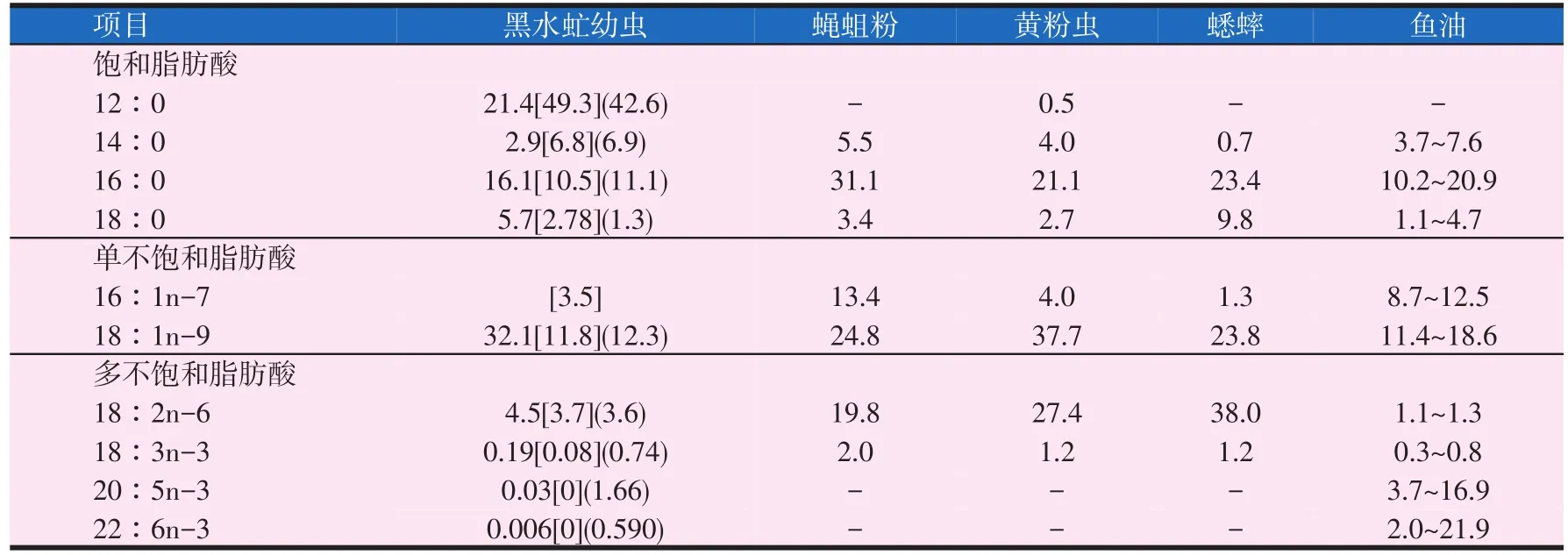
表3 昆虫脂肪酸组成(%)
1.3 碳水化合物
昆虫体内含有1%~10%左右的糖类,包括葡萄糖、果糖、糖原及存在于血液中的海藻糖。昆虫体表还含有大量以乙酞氨基葡萄糖为主要成分的几丁质(甲壳素),一般含量为5%~15%。不同虫态的昆虫几丁质含量也不同,如家蚕(Bombyxmori Linnaeus)干蛹几丁质含量为3.73%,脱脂蛹为5.55%。云南松毛虫(DendrolimushouiLajonquiere)蛹为7.47%,成虫可高达17.83%。虫茶为昆虫排泄物的一种,糖类含量也较高,可达16.27%。
1.4 矿物质
昆虫含有丰富的矿物质,如K、Na、Ca、Cu、Fe、Zn、Mn、P等,许多食用昆虫的Ca、Zn、Fe、Mg、K等含量较高,柞蚕含较高含量的K、Mg,家蝇含较高的K、Mg、P等矿物元素。黄粉虫含有较多的P、K、Fe、Na和Ca等多种元素。蚯蚓体内含有多种常量、微量元素(如Mn、Zn、Ca、Cu、Mg、Fe、Na、K、Se),Fe含量比鱼粉高13倍,Mn含量比鱼粉高5倍,Zn含量比鱼粉高3倍,Cu含量比鱼粉高1倍,其中P的利用率高达90%以上。中华稻蝗含有Na、Mg、Fe、Zn、Se等9种元素;蚂蚁含有丰富的Mn、Zn、Se、Mg、Ca、P、Fe等28种人体所必需的元素,其含Zn量比大豆高8倍,比猪高2倍(孙红梅,2011)。
2 昆虫资源饲料化利用的方式
2.1 整体直接利用
昆虫资源作为一种优质饲料原料,在水产饲料研发中已有一定的报道。目前研发较多的有蚕蛹、黑水虻、黄粉虫等,多数为整体直接利用。以消化率为主的研究表明,蚕蛹可完全或部分替代黄鳝(Monopterus albus)饲料中的鱼粉(刘丹丹等,2009)。在框鲤(Cypriniformes)饲料中蚕蛹添加水平为5.5%,可替代50%鱼粉蛋白(吉红等,2012)。胡俊茹等(2014)发现在黄颡鱼(Pelteobagrus fulvidraco)幼鱼饲料(粗蛋白41%,粗脂肪7.6%)中使用黑水虻幼虫粉可替代鱼粉用量的30%而不影响生长,并显著降低血清谷丙转氨酶、谷草转氨酶、尿素氨以及甘油三酯的含量。黑水虻虫体或干粉替代鱼粉、豆粕等蛋白源,饲养包括斑点叉尾鮰(Iczalurus punczarus)、罗非鱼(Oreochromis aureus)、虹鳟鱼(Oncorhynchus mykiss)等(St-Hilaire,2007b),均取得了较好的养殖效果。用6%~8%的鲜黄粉虫喂养甲鱼、鳗鱼等水产动物,由于其具有可口性好、助消化的特点,水产动物长势快、抗病力强。文远红等(2013)发现,用蝇蛆粉替代鱼粉饲喂黄颡鱼幼鱼,其用量不宜超过20%,否则会对其生长及部分血浆生化指标产生影响。Haro等(2015)发现在金头鲷(Sparusaurata)日粮中添加不同水平的丝光绿蝇幼虫可提高鱼体肌肉组织中花生四烯酸的含量,其营养价值可同鱼粉等饲料原料媲美。Ngoc等(2016)则报道用不同水平的蚯蚓粉(30%、70%、100%)分别替代鱼粉饲喂鲤鱼(Cyprinus carpio),生长效果均比较理想。
2.2 提脂后进行利用
将昆虫体蛋白质和脂肪进行分离后利用,也是一种较好的方法。可以使用冷榨法脱去昆虫粉中的脂肪,还可参照油料作物的提脂方法进行昆虫脂肪的提取。正己烷可有效地将蚕蛹脱脂、脱色、除臭,且提高了蚕蛹的粗蛋白含量(吉红,2016)。研究表明,脱脂蛆粉可以提高蛆粉在日粮中的添加水平而不影响非洲鲶鱼的生长性能(Fasakin等,2003);然而,含有高水平高温烘干法制作的脱脂黑水虻粉的日粮与含有低水平常温烘干法制作的脱脂黑水虻相比,抑制了太平洋鲑鱼(Oncorhynchus spp)的生长(Lock等,2014)。
2.3 发酵或酶解利用
对昆虫原料进行发酵或酶解,可将大分子物质转成小肽等活性物质。李峰等(2014)用纳豆菌发酵黄粉虫,制备出具有调节脂质代谢、抗氧化、增强抵抗力等能力的低分子蛋白肽,肽得率高达59%。在发酵过程中,细菌会把饲料成分变得更易消化,并把糖转换成酸,降低pH值,抑制腐败菌的繁殖。Rangacharyulu等(2003)分别用新鲜蚕蛹和发酵蚕蛹完全替代饲料中的鱼粉(对照组饲料中鱼粉添加水平为4.5%),对池塘混养的卡特拉鲃(Catlacatla)、印度鲮(Cirrhinusmrigala)、南亚野鲮(Labeorohita)和白鲢(Hypophthalmichthys molitrix)进行投喂,发现新鲜蚕蛹组和鱼粉组之间各项指标没有显著差异,而发酵蚕蛹组在增重率、饲料系数和特定生长率方面均显著优于新鲜蚕蛹组和鱼粉组。发酵蚕蛹替代饲料中25%的鱼粉蛋白对框鲤幼鱼生长、饲料利用和体成分无负面影响,且有降低试验鱼血脂,保护肝胰脏的作用,但更高水平的替代则会抑制框鲤幼鱼生长,降低饲料利用率,危害其健康状态(Zhou等,2016)。
张莉莎等(2009)采用胰蛋白酶及碱性蛋白酶共同水解黄粉虫,发现获取的低分子肽具有较高的水解度,酸溶性肽得率高达91%。许彦腾(2015)对脱脂黑水虻进行酶解,产物中蛋白含量高达91%,营养丰富,质量优于脱脂黑水虻粉,具有一定的抗氧化性。邓方坤(2015)开发出了一种给昆虫喂食昆虫病毒,利用病毒产生的蛋白水解酶裂解昆虫虫体,从而提取昆虫多肽的方法。此外,酸水解法可增加蚕蛹和蛆粉蛋白质含量而减少其脂质量(Fasakin等,2003),但可能会破坏一些必需氨基酸例如胱氨酸、蛋氨酸和色氨酸。另外,用糖蜜、酸和抗氧化剂青贮昆虫粉是比较好的处理方法(Newton等,2005)。
3 昆虫资源饲料化研究实例
3.1 蚕蛹
Ji等(2015)发现,蚕蛹等蛋白替代超过50%鱼粉蛋白时,建鲤(CyprinuscarpioVar.Jian)幼鱼生长性能显著下降,50%替代组肌肉粗蛋白含量显著高于80%替代组。脱脂蚕蛹替代建鲤幼鱼饲料鱼粉不应超过50%,同时添加晶体氨基酸可提高鱼体的蛋白沉积水平(张建禄等,2013)。随着脱脂蚕蛹替代水平升高,建鲤肠道菌群多样性下降,而饲料中补充赖氨酸后这种影响会减弱(钟雷等,2014)。蚕蛹替代吉富罗非鱼(Oreochromisniloticus)日粮中50%鱼粉可提高鱼体生长性能,促进鱼体蛋白质沉积,提高血清溶菌酶活力,且可降低肝脏和血液中脂肪含量(王淑雯等,2016)。日本鲈鱼(L.japonicus)对未脱脂蚕蛹粉的能量消化率低于对禽副产物粉、羽毛粉、血粉和豆粕的消化率,和肉骨粉消化率接近,另外,粗蛋白消化率也低于禽副产物粉、血粉和豆粕,和羽毛粉类似,高于肉骨粉(Wen等,2010)。
3.2 蝇蛆和蝇蛹
非洲鲶鱼(Clarias gariepinus)日粮中蝇蛆粉添加水平应在25%~30%,添加水平超过30%后鱼体生长性能有下降趋势(Madu等,2003)。用麦麸∶活蝇蛆=4∶1的混合物饲喂尼罗罗非鱼(Tilapianilotica),鱼体生长性能、饲料转化率和存活率均优于全麦麸对照组(Ebenso等,2003)。尼罗罗非鱼日粮中可以添加15%~68%蛆粉,25%添加水平(替代34%的鱼粉)显示生长性能和存活率最好,对血液学指数和内环境稳态没有负面影响。然而,适当的多不饱和脂肪酸源添加到日粮中,以改善鱼体脂肪酸组成(Ogunji等,2007、2008)。
3.3 黄粉虫
Gasco等(2014a)认为,虹鳟(Oncorhynchus mykiss)日粮中黄粉虫幼虫可添加至50%,Belforti等(2015)则发现,日粮添加50%黄粉虫,虹鳟摄食率显著低于对照组,表观消化率也显著低于其余各组;日粮添加25%和50%黄粉虫,饲料系数显著低于对照组,鱼体成活率显著高于对照组;肌肉蛋白含量随着替代水平升高而升高,而脂肪含量降低。黄粉虫替代60%的日粮鱼粉(26%添加水平)对非洲鲶鱼(Clariasgariepinus)生长性能和饲料利用率没有显著影响,但是添加水平提高至35%~43%(替代鱼粉80%~100%)时,鱼体生长性能和饲料利用率和蛋白质效率会降低(Ng等,2001),而Roncarati等(2015)最新的研究则表明,日粮添加黄粉虫粉会降低鲶鱼(Silurusasotus)幼鱼增重。在鲷(Dicentrarchuslabrax)、欧洲海鲈(Sparusaurata)日粮中黄粉虫替代25%的鱼粉不会显著影响鱼体生长,而替代50%鱼粉时,生长性能显著降低,且欧洲海鲈肌肉中n-3HUFA水平下降(Gasco等,2014b;Piccolo 等,2014)。
3.4 黑水虻
黑水虻幼虫可以部分或者完全替代某些养殖鱼类日粮中的鱼粉。大西洋鲑(Salmo salar)日粮中添加5%~25%的黑水虻粉,并添加赖氨酸和蛋氨酸,试验鱼的体增重未受影响(Lock等,2014)。而日粮中添加12%~30%的黑水虻粉显著降低斑点叉尾鮰、虹鳟和大菱鲆(Scophthalmus maximus)的生长性能(Newton等,2005;St-Hilaire等,2007b;Kroeckel等,2012)。幼年大菱鲆日粮中黑水虻预蛹粉添加水平可以达到33%,且对摄食量和饲料转化率没有显著性影响,但特定生长率显著降低;添加水平高于33%时,会造成饲料适口性和蛋白质消化率下降,引起大菱鲆摄食量下降和生长性能下降(Kroeckel等,2012)。日粮中添加25 g/kg黑水虻油对建鲤幼鱼生长性能、饲料利用率和血清生化指标没有负面影响,且提高了鱼体肌肉中n-3PUFA的水平,降低了n-6PUFA的水平。黑水虻油添加水平大于等于18.75 g/kg时,建鲤腹脂指数显著降低(Li等,2016)。
3.5 蝗虫
沙漠蝗虫粉可以替代尖齿胡鲶(Teugels)日粮中25%的日粮蛋白,而不会降低其生长性能,当替代水平高于25%时,则造成生长性能和饲料效率下降,这可能与昆虫粉中的几丁质有关(Balogun,2011)。臭腹腺蝗粉可以等量替代尖齿胡鲶幼鱼日粮中25%的鱼粉,而不会降低其生长性能和营养利用率,而高添加水平会降低鱼体消化率和生长性能(Alegbeleye等,2012)。91 d的饲养试验表明,胡子鲇(Clariasbatrachus Linn)日粮中添加蝗虫粉对鱼体血液学参数没有影响,但会造成鳃的轻微皱缩和卵巢类固醇水平降低,这可能对其繁殖有影响(Johri等,2011)。
4 昆虫资源饲料化开发存在的营养饲料学问题及对策
4.1 生物富集有毒物质
杀虫剂、重金属和天然毒素等都有可能通过养殖基质富集在昆虫体内。因此,昆虫养殖机构需优化养殖基质,以有效控制昆虫体内富集的毒素等有害物质(Spiegel等,2013)。在有机质副产品作为昆虫养殖基质的情况下,控制养殖基质更为重要。用家禽粪便饲养获得的蝇蛆幼虫加工成蛆粉,添加到尼罗罗非鱼日粮中饲喂罗非鱼(Tilapia nilotica)后,发现鱼体肝脏谷胱甘肽转移酶水平升高,这可能与残留在鸡粪便中的农药、医用药品或有毒物质有关(Ogunji等,2007)。
4.2 氨基酸组成不平衡
植物蛋白源替代鱼粉的研究发现,使用混合日粮有助于预防使用单一原料造成的鱼体氨基酸缺乏(Hu等,2013)。同样,使用混合昆虫粉可以避免单一来源昆虫粉造成的日粮氨基酸不平衡问题。例如黑水虻粉缺乏色氨酸,而家蝇粉和蚕蛹粉富含色氨酸,则可将黑水虻粉与家蝇粉或者蚕蛹粉混合使用。昆虫粉也能和其它来源的蛋白质混合,研究表明大麦虫和10%益生伞菌混合添加到鱼饲料中,能改善罗非鱼(Oreochromis sp.)的生长性能(改善特定生长率、饲料转化率、蛋白效率和存活率)(Din等,2012)。作者认为这可能是因为这种方式平衡了日粮氨基酸组成或者是伞菌起到了益生作用。
4.3 必需脂肪酸缺乏
通常认为陆生昆虫粉添加到海水鱼饲料中的限制性因素为HUFA缺乏,而n-3和n-6脂肪酸可以通过调控昆虫养殖基质来提高(Ogunji等,2008)。饲喂牛粪便的黑水虻幼虫脂肪主要含饱和脂肪酸、单不饱和脂肪酸和微量的n-3脂肪酸(0.2%),而饲喂78%牛粪和22%鱼粪混合物的幼虫,24 h后n-3脂肪酸的含量就能达到4%(St-Hilaire等,2007a)。用含有鱼粪(25%或50%)的基质饲喂黑水虻幼虫一个月,在虹鳟日粮中黑水虻幼虫替代鱼粉比例就可以提高25%~50%,而不影响试验鱼的生长性能(Sealey等,2011)。由于水生昆虫捕食性比草食或杂食陆生昆虫强,且它们摄食的水生微生物富含n-3脂肪酸,因此水生昆虫n-3HUFA(主要是EPA)含量比陆生昆虫高(Fontaneto等,2011),更适合添加到海水鱼饲料中。
4.4 几丁质影响营养物质的消化吸收
研究发现,某些种类的昆虫粉以一定比例添加到鱼类饲料中,会导致鱼类生长性能下降,以及蛋白质和脂质消化率降低。一般认为这是由其所含的几丁质造成的,但这方面尚无确凿的证据(Alegbeleye等,2012)。消化几丁质需要3种酶,几丁质酶、壳二糖酶和溶菌酶,这些酶在肉食性和杂食性鱼类消化系统中都有发现。在鱼胃中发现的几丁质酶可以破坏几丁质外骨骼。在鱼的肠道中发现的壳二糖酶与营养吸收有关。这些酶在淡水鱼和海水鱼都存在。但用黑水虻幼虫饲喂大菱鲆,检测了鱼中肠,未发现任何几丁质酶活性(Kroeckel等,2012)。
几丁质包裹在由蛋白质和矿物质(大部分是钙)构成的网络里,昆虫甲小皮是由包裹在蛋白质、脂质和其它混合物中的几丁质组成的。这些包裹几丁质的网络,可以降低几丁质酶或者蛋白酶接触几丁质,并阻止肠道对蛋白质和脂质的吸收,这样不仅降低几丁质的消化率,而且也降低了脂质和蛋白质的消化率,从而造成鱼类对饲料营养的利用率也降低,其生长性能也下降。用含有2%、5%和10%几丁质的日粮饲喂杂交罗非鱼(尼罗×奥利亚)时,出现同样的情况。然而也有与此相反的研究结果,鱼类或者不受日粮几丁质影响,或者生长性能提高。军曹鱼(Rachycentroncanadum)饲喂包含3%的几丁质的蟹粉或者包含10%几丁质的虾粉,蛋白质消化率升高(Fines等,2010);1%的日粮几丁质不影响鲤鱼的生长性能(Gopalakannan等,2006);5%和10%日粮几丁质的刺激真鲷(Pagrus major)的生长,然而20%的几丁质降低真鲷的生长。另外,有研究表明低水平的日粮几丁质刺激海鲤(Sparusaurata L.)(<0.01%)(Esteban等,2001)和鲤鱼(<1%)(Gopalakannan等,2006)内源性免疫系统的活性,并且增加鲤鱼对细菌性疾病的抵抗力。
通常认为单胃动物,包括鱼,没有消化几丁质的能力。通过碱性提取法(Belluco等,2013),可以除掉昆虫粉中的几丁质,从而可以将昆虫粉加入水产饲料中。几丁质酶或几丁质消化菌可以添加到昆虫粉中以提高鱼类对几丁质蛋白复合物的消化率(Kroeckel等,2012)。可以通过化学或者酶解方法处理几丁质,将其分解为甲壳低聚糖(COS)、乙酰氨基葡萄糖(GlcNAc)或者壳聚糖。然而,这些加工工艺提高了昆虫粉的成本。
4.5 适口性差
昆虫粉的适口性差,这可能是由化学或微生物污染、抗营养因子、黄酮类似物和萜烯类化合物等多种因素造成的。陆生昆虫富含单不饱和脂肪酸(Barroso等,2014),易于氧化酸败,从而导致适口性变差。干燥、水解、青贮或者脱脂加工可改善昆虫粉的适口性、营养利用率、消化率等。研究表明,直接晒干或烘干,或者蒸煮后晒干或烘干,可以改进昆虫粉的适口性。通常认为烘干比晒干好,晒干后的昆虫粉富含油脂(会增加脂质氧化的风险),而蛋白质含量较少(Aniebo等,2010)。
5 昆虫作为水产饲料原料的产业化前景
昆虫产品作为饲料原料,具有品质优良、营养全面及成本低廉的优点。据测算,大约2 kg生物废弃物就能生产1 kg昆虫生物质。因此,将昆虫资源应用于水产饲料生产,可降低水产养殖对豆粕、鱼粉等饲料资源的消耗,从而避免养殖业对南美鱼粉和豆粕的过度依赖。
另一方面,昆虫资源开发作为一种新兴的产业,其资源饲料化利用依然存在一些问题,如资源状况不够稳定,这首先表现为量的不稳定。昆虫资源要用作饲料原料,首先需要规模化供应,由于昆虫生产对饲养环境(温度和湿度)、群体等的要求比较严苛,尤其在繁殖时期更是如此,因此规模化养殖仍有技术瓶颈未能突破。其次是产品品质不稳定。由于生产流程尚未标准化,以及利用成分不稳定的餐厨垃圾和畜禽粪便生产昆虫,昆虫产品品质的稳定性就得不到保障。即使一些开发历史较长的昆虫资源,如蝇蛆、黄粉虫、黑水虻幼虫等,也由于缺乏营养、加工、饲养等方面的系统研究,产业化方面存在诸多问题需要解决。
再者,从产业链的角度分析,昆虫的深加工可大大提高其附加值和利用率,如采取热机械处理、研磨、挤压、超临界CO2抽提、烘干等工艺,把昆虫蛋白和脂肪分离开进行使用,可更加精准地发挥昆虫资源的优势,其产品甚至可在其他领域得以利用,例如从黑水虻或者黄粉虫中分离出的油脂可以用作生物燃料的原料(Stamer,2015)。而目前研究主要关注于昆虫的营养素含量和营养特性(Makkar等,2014),对于昆虫蛋白质或脂肪的物理化学特性或功能特性的关注偏少。已有研究表明,加工、制备或贮藏过程会对昆虫蛋白的物理化学和功能特性产生影响(Azagoh等,2016)。
今后,应采取产学研紧密结合的路径,深入开展科学研究,突破昆虫养殖技术瓶颈,推动昆虫资源规模化生产,并关注加工过程对昆虫成分物理化学和功能特性的影响,提升产品品质,进一步发挥昆虫资源的作用,使其应用价值最大化。总之,昆虫资源的开发利用前景非常广阔,相关工作的深入将为未来水产业的可持续发展提供重要支撑。
猜你喜欢
作文小学中年级(2021年3期)2021-12-25
食品工业科技(2021年22期)2021-11-14
作文·小学低年级(2021年3期)2021-11-02
环境卫生工程(2021年2期)2021-06-09
生物技术通报(2021年4期)2021-05-14
绿色科技(2019年10期)2019-06-17
广西林业科学(2019年1期)2019-04-26
中国酿造(2017年8期)2017-09-03
小溪流(画刊)(2017年6期)2017-07-10
小天使·四年级语数英综合(2017年4期)2017-04-18
