
调水调沙后黄河口邻近海域浮游植物群落响应特征
2015-06-24 14:10:16苏芝娟王玉珏董志军张义文刘东艳王艳霞
海洋学报 2015年4期
苏芝娟,王玉珏,董志军,张义文,刘东艳*,王艳霞
(1.河北师范大学 资源与环境科学学院 河北省环境演变与生态建设省级重点实验室,河北 石家庄 050024;2.中国科学院烟台海岸带研究所 中国科学院和山东省海岸带环境过程重点实验室,山东 烟台 264003;3.河北省科学院地理科学研究所,河北 石家庄 050021)
调水调沙后黄河口邻近海域浮游植物群落响应特征
苏芝娟1,2,王玉珏2,董志军2,张义文1,刘东艳2*,王艳霞3
(1.河北师范大学 资源与环境科学学院 河北省环境演变与生态建设省级重点实验室,河北 石家庄 050024;2.中国科学院烟台海岸带研究所 中国科学院和山东省海岸带环境过程重点实验室,山东 烟台 264003;3.河北省科学院地理科学研究所,河北 石家庄 050021)
为研究黄河调水调沙对邻近海域浮游植物群落的影响,2013年7月在黄河第16次调水调沙事件后,开展了水文、化学与生物综合调查。研究结果表明,黄河口邻近海域温度、营养盐浓度整体呈现由河口向离岸区域逐渐递减的分布趋势,盐度呈现由河口向离岸区域逐渐递增的分布趋势,显示了黄河水输入的影响程度。叶绿素a与营养盐浓度在空间分布上呈现出较好的对应关系,在黄河口偏渤海湾侧明显高于偏莱州湾侧,且近河口区明显高于离岸区。营养盐结构分析表明,黄河口邻近海域普遍存在磷酸盐(DIP)的绝对和相对限制;但黄河水沙输入在局部站位缓解了硅(DSi)限制。浮游植物群落结构的空间变化显著受到盐度的影响,在受黄河水输入影响显著的C、D、E断面,蓝藻与绿藻的生物量比例明显增高;影响相对较弱的断面则以硅藻、甲藻为主。浮游植物群落结构与环境因子的主成分分析结果表明,DSi、DIP和盐度是影响该海域浮游植物空间变化的关键环境因子。甲藻、蓝藻与绿藻群落受盐度变化的影响程度明显大于硅藻群落;但甲藻群落对营养盐结构的敏感性低于硅藻、蓝藻和绿藻群落。
黄河口;浮游植物;调水调沙;盐度;营养盐
1 引言
河流水沙入海对河口三角洲及邻近海域生态系统的物质循环与环境演变起到重要作用,例如,埃及尼罗河、美国科罗拉多河、密西西比河、南美亚马孙河、中国的长江、黄河[1—3]。然而,近几十年来,由于气候变化以及水坝建设、引水引沙、土地利用等人为因素,入海径流量和输沙量显著下降,已经引起区域甚至全球尺度上的河口、三角洲及其近岸生态环境变化。典型的例子如尼罗河三角洲,在入海径流与泥沙减少后,三角洲盐度快速升高、生源要素输入减少,导致浮游植物生物量减少了95%,渔获量减少了80%[4]。黄河作为世界第五大河,每年约有数亿吨泥沙入海,这些高通量的泥沙输运伴随着年均约1.4万吨的营养盐入海,对黄河三角洲与莱州湾邻近海域的环境演变与生态系统发育构成显著影响[5]。已有研究表明,黄河改道与水沙输入变化已经引起三角洲岸线的变迁,邻近海域环流场的变化,并影响到渔业资源[3,6]。
2002年以来,黄河水利委员会定期对黄河实施调水调沙,其径流量在短时间内激增,可对黄河口邻近海域环境因子乃至生态系统产生显著影响[7—8]。浮游植物和浮游动物分别是海洋中重要的生产者和消费者,对海洋环境反映敏感,海洋中的温盐结构和营养盐浓度变化可显著影响浮游生物群落结构变化[9—10]。然而,关于黄河调水调沙对邻近海域生态系统影响的研究则相对较少。已有研究发现,调水调沙对黄河口动物群落结构可产生一定程度的影响,但不是导致群落更替的关键因素[11—12];而对浮游植物群落结构的影响则十分显著[13]。然而,调查资料很有限,仅限于2009年7月的网采调查,且缺乏与关键环境因子的相关性分析,难以综合浮游植物群落空间格局变化的响应特征。因此,本研究在2013年7月黄河第16次调水调沙事件后,对黄河口及其邻近海域开展了水文、化学、生物的综合调查。通过对该海域浮游植物群落结构、温盐结构以及营养盐空间变化特征的研究以及相关性分析,来阐明黄河调水调沙后浮游植物群落结构与环境因子的相应关系。研究结果为进一步了解黄河调水调沙事件的生态影响提供参考依据。
2 材料与方法
2.1 调查站位
2013年7月26—30日期间,对黄河口及邻近水域(37.40°~38.11°N,119.06°~119.69°E)开展了水文、化学与生物的综合调查。依据黄河冲淡水的扩散特征,从河口到离岸海域共设置6个断面(A~F)、29个站位(图1)。调查区域平均水深11 m,最浅处4 m,最深处18 m左右。

图1 黄河口邻近海域采样站位(a)与水深(m)(b)Fig. 1 Sampling sites (a) and water depths (m)(b) in adjacent sea of the Yellow River mouth
2.2 样品采集与分析
调查区域海水的温度和盐度数据用YSI6920型多参数水质监测仪测定。用5 L的卡盖式采水器(国家海洋技术中心)采集表层水样分别用于营养盐、叶绿素a和浮游植物分析。取500 mL海水装入聚乙烯瓶,并立即加入福尔马林溶液固定(浓度为3%~5%),用于浮游植物种类鉴定与数量计数。取1 000 mL水样经GF/F滤膜(Whatman)过滤后,滤膜于-20℃下避光并冷冻保存,用于叶绿素a浓度的测定。每个采样点取3个平行样,结果取其平均值。另取250 mL水样用0.45 μm醋酸纤维滤膜滤后,滤液用于营养盐浓度的测定。
营养盐参数利用德国Bran+Luebbe AA3营养盐自动分析仪测定,包括溶解无机氮(DIN:dissolved inogranic nitrogen,为NO3-N、NO2-N、NH4-N之和)、磷酸盐(DIP:dissolved inorganic phosphate,为PO4-P)和硅酸盐(DSi:dissolved inorganic silicate,为SiO3-Si)。其中,硝酸盐(NO3-N)采用镉-铜还原后再用盐酸-萘乙二胺比色法测定,检出限为0.03 μmol/L;亚硝酸盐(NO2-N)采用盐酸-萘乙二胺比色法测定,检出限为0.005 μmol/L;氨氮(NH4-N)采用水杨酸钠法测定,检出限为0.04 μmol/L;磷酸盐(PO4-P)采用磷钼蓝法,检出限为0.02 μmol/L;硅酸盐(SiO3-Si)采用硅钼蓝法,检出限为0.03 μmol/L。
叶绿素a浓度采用分光光度法测定[14]。保存叶绿素a的滤膜先用90%的丙酮在4℃ 避光条件下萃取24 h,在4 000 r/min转速下离心10 min后,用紫外可见分光光度计(TU-1810),在波长750 nm、664 nm、647 nm与630 nm处分别测定溶液吸光值(90%丙酮作空白),将所得数据用以下公式换算成叶绿素a的浓度CChl a(单位:μg/L):
(1)
式中,E750、E664、E647与E630分别是各波长下的吸光值,V丙酮为丙酮萃取液的体积(单位:mL);V水样为过滤海水的体积(单位:L)。
浮游植物计数和鉴定在Olympus IX 81型倒置光学显微镜200~400倍下进行。浮游植物计数采用Utermöhl方法[15],样品充分摇匀后取出25 mL放入Utermöhl计数框,静置24 h,进行浮游植物的种类鉴定和计数。对于高浊度样品,为避免太多颗粒物的影响,沉降时间稍短一些。浮游植物种类的鉴定参考书为《中国海藻志》、《海藻学概论》、《海藻学》、《中国近海赤潮生物图谱》、《中国海域常见浮游硅藻图谱》、《中国海域甲藻扫描电镜图谱》、《中国海域甲藻Ⅰ》等[16-22]。
2.3 数据处理
物种多样性指数采用香农-威纳指数(H′,Shannon-Wiener index)[23],其计算公式为:
(2)
物种均匀度(J)采用Pielou 的计算公式[24]:
(3)
式中,S为样品中总种类数,pi为第i种的个体数与样品中的总个数的比值。
站位图、营养盐及生物多样性指数的平面分布图采用Surfer 8.0软件绘制;群落结构与环境因子的相关性采用主成份分析方法,数据经过lg(x+1) 转换后,使用Canoco for Windows 4.5软件绘制群落结构与环境因子的主成分分析排序图。
3 结果与讨论
3.1 表层海水温度和盐度的空间分布特征
图2为调查海区各断面温度与盐度分布特征。结果显示:调查期间,表层海水温度范围为22.9~27.7℃,平均值为25.2℃(见图2a);盐度范围为15.5~27.5,平均值仅为20.8(见图2b),显示大量淡水输入的影响。海水温度的空间变化并不明显,随着水深的增加,站位间的温度差异减小。仅F断面显示了较高的水温特征,范围为26.3~27.7℃,均值为27℃;可能与F断面的水深较浅有关。比较而言,盐度则显示了显著的空间变化,在各断面从近岸向外海逐渐呈上升趋势,显示了淡水输入影响逐渐减弱。其中,离河口最近的C断面和D断面盐度最低。C断面的盐度介于16.8~23.2,平均值为18.5;D断面介于17.6~21.7,平均值为19.2。而离河口最远的F断面,盐度相对最高,介于21.1~24.6,平均值为22.7。温度与盐度的空间变化响应了黄河冲淡水的影响。
3.2 表层海水营养盐的空间分布特征
调查期间,各断面营养盐浓度与结构的空间特征如图3所示。结果显示:DIN、DIP与DSi三种营养盐浓度均呈现出由河口向东北离岸区域递减的空间特征。其中,DIN浓度平均值为41.6 μmol/L,变化范围为17.2~69.6 μmol/L;在黄河口靠近渤海湾侧为高浓度区,最高值出现在B1—B2站,分别为69.5 μmol/L与69.6 μmol/L;最低值出现在B5站,浓度为17.2 μmol/L(见图3a)。DIP浓度平均值为0.09 μmol/L,变化范围为0.01~0.19 μmol/L;最高值出现在B1站,浓度为0.19 μmol/L;最低值出现在B5站,浓度为0.01 μmol/L(见图3b)。DSi浓度平均值为39.6 μmol/L,变化范围为4.8~91.3 μmol/L;其浓度的空间变化趋势与DIP相似,最高值亦出现在B1站,浓度为91.3 μmol/L;最低值出现在B5站,浓度为4.8 μmol/L(见图3c)。营养盐浓度的空间分布特征,与盐度的变化梯度基本一致,显示黄河淡水输入的影响随水深逐渐减弱的空间特征。本次调查结果与2010和2011年枯水期的调查结果相比[25],DIN和DIP浓度大幅度降低,DSi浓度大幅度增高。这与已有研究结果一致,即DIN和DIP浓度丰水期低于枯水期,DSi浓度夏季高于其他季节[7,26]。此外,夏季,水体从近渤海湾湾顶处向南构成反时针环流[27],影响到物质向外海的输入,这可能是黄河口靠近渤海湾侧营养盐浓度相对较高的重要原因[28—29]。
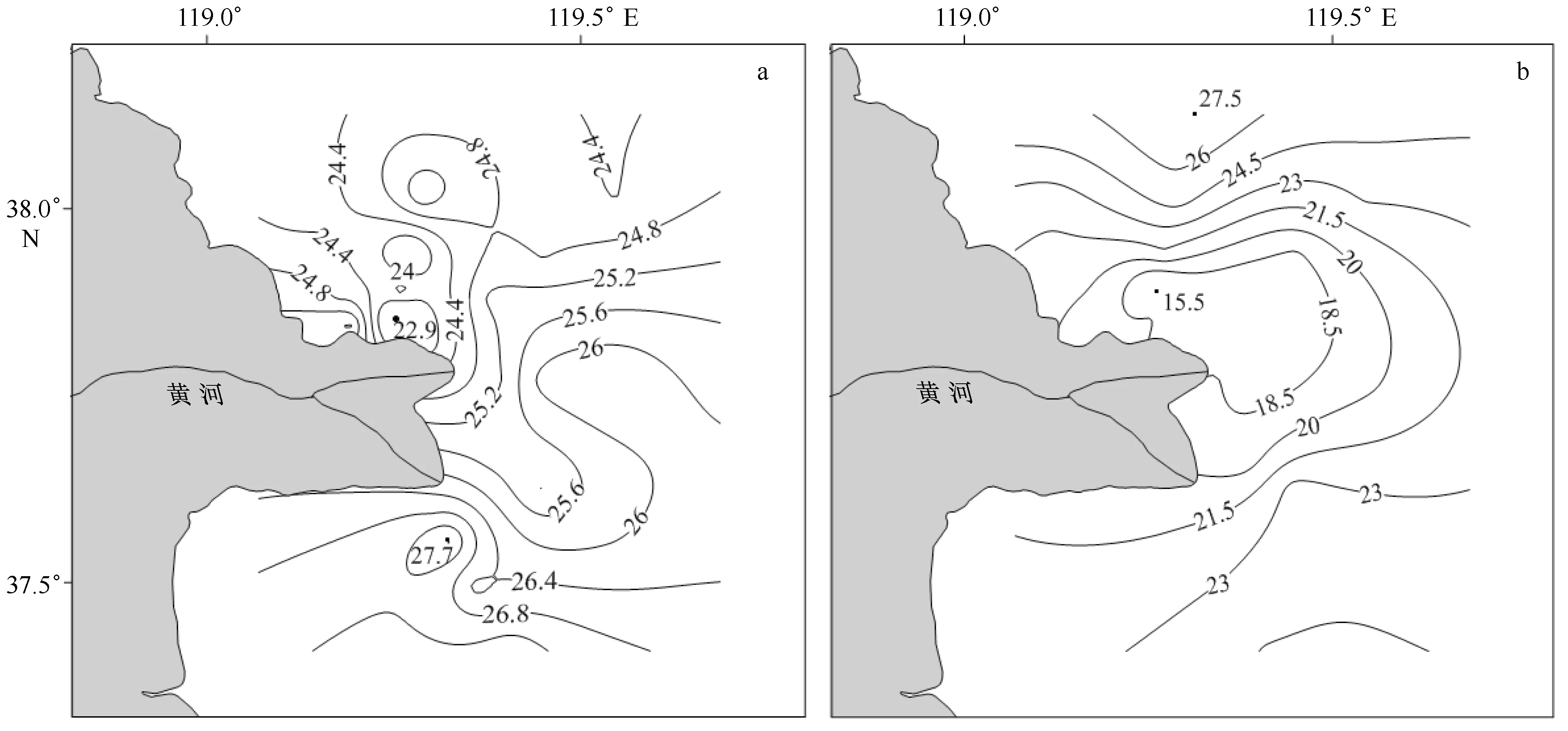
图2 调查海区表层海水温度(℃)(a)和盐度(b)的空间分布特征Fig.2 The spatial distribution of surface seawater temperature (℃)(a) and salinity (b) during the survey
营养盐结构变化对浮游植物群落结构有着显著影响[30—31]。以往研究结果显示,近40年来,渤海水域营养盐结构发生了明显变化,N/P比例严重失调,磷和硅作为限制因子出现的几率明显增加[32—35]。根据 Nelson和Brzezinski[36]、Justic 等[37]提出的浮游植物生长的化学计量和可能营养盐限制因素标准,DIN、PO4和SiO3的浓度分别低于1 μmol/L、0.1 μmol/L和2 μmol/L时,对浮游植物生长造成绝对限制;N/P比小于10,Si/N比大于1时,DIN存在相对限制;N/P比和Si/P比大于22时,PO4出现相对限制;Si/N比小于1,Si/P比小于10时,SiO3存在相对限制。
本次调查结果显示:DIN/DIP的变化范围为217.2~1 562.9,平均值为592.7,分布趋势为由河口向离岸递增,出现了明显的磷酸盐限制现象;尤其在F断面,出现了磷酸盐的相对与绝对限制(见图3d)。DSi/DIN的变化范围为0.2~1.8,平均值为0.9;分布趋势与DIN/DIP正好相反,高值区分布在河口,由河口向东北离岸递减,说明河口输入对硅酸盐的补充,显著缓解了该区域的硅限制状态(见图3e)。最高值出现在C1站位,比值为1.8;低值区分布在河口南部的F断面,最低值出现在F2站位,比值仅为0.2,表明仍存在硅酸盐的限制(见图3e)。DSi/DIP的变化范围为267.4~526.3,平均值为409.0;其分布趋势有两种,一是由河口向东北离岸递减,研究区最高值就出现在离河口较近的B2站,比值为526.3。二是研究区南部出现了一个低值中心,并向东南部离岸递增。最低值出现在F2站,比值为267.4(见图3f)。
通过对营养盐限制因子出现几率进行计算发现:DIN与DSi浓度分别低于1 μmol/L和2 μmol/L的出现几率为0;DIP浓度低于0.1 μmol/L出现几率为62.1%。N/P比小于10,Si/N比大于1时,出现几率为0;Si/N比小于1,Si/P比小于10时,出现几率为0;N/P比和Si/P比大于22时,出现几率为100%。这一结果表明黄河径流量的增加显著提高无机氮与硅酸盐的输入量,但对磷酸盐的输入贡献相对不大。
3.3 浮游植物群落结构的空间分布特征
本次调查,共鉴定浮游植物5门60属100种(见附表1),主要以硅藻和甲藻为主,包括硅藻63种,占总物种数的63%;甲藻24种,占总物种数的24%;蓝藻8种,占总物种数的8%;绿藻3种,占总物种数的3%;金藻1种,占总物种数的1%;还有未定类的三深碎裂藻,占总物种数的1%。物种组成显示了大河口盐度低,物种多样性高,半咸水物种多的特点[38—39]。这与黄河口历史调查结果基本一致,例如,田家怡[40]以1996年3、5、8、11月份的定量调查资料,鉴定出该海域浮游植物72种,分别隶属于硅藻门、甲藻门、金藻门、绿藻门;冷宇等[41]对该海域2004—2010年每年夏季的调查共获得浮游植物137种,包括硅藻门、甲藻门、金藻门、黄藻门;刘晓彤等[13]对该海域2009年夏季的调查,共鉴定浮游植物29属53种,包括硅藻门、甲藻门、蓝藻门、金藻门、绿藻门。

图3 营养盐浓度(μmol/L)与结构的空间分布特征:a.DIN,b.DIP,c.DSi,d.DIN/DIP,e.DSi/DIN,f.DSi/DIPFig.3 The spatial distribution of nutrient concentrations (μmol/L) and structures during the survey: a.DIN,b.DIP,c.DSi,d.DIN/DIP,e.DSi/DIN,f.DSi/DIP
调查期间,海水叶绿素a浓度范围为1.1~32.7 μg/L,平均值为6.4 μg/L(图4a)。叶绿素a浓度的空间分布与营养盐浓度的空间分布很接近。在靠渤海湾侧出现一个高值区,最高值出现在A1站位,为32.7 μg/L。这与已有研究结果一致,1989年夏季该海域浮游植物细胞丰度在靠近渤海湾侧的近岸出现高值区,并呈弧形由近岸到远岸逐渐降低[42]。浮游植物群落的香农-威纳多样性指数(H′)和Pielou均匀度指数(J)的分布趋势都是在研究区的中部站位出现高值和西北部高,而靠近岸和外海的站位出现较低值(图4b,4c)。其中,香农-威纳多样性指数介于0.39~4.46,平均值为3.16;Pielou均匀度指数介于0.16~0.91,平均值为0.75。
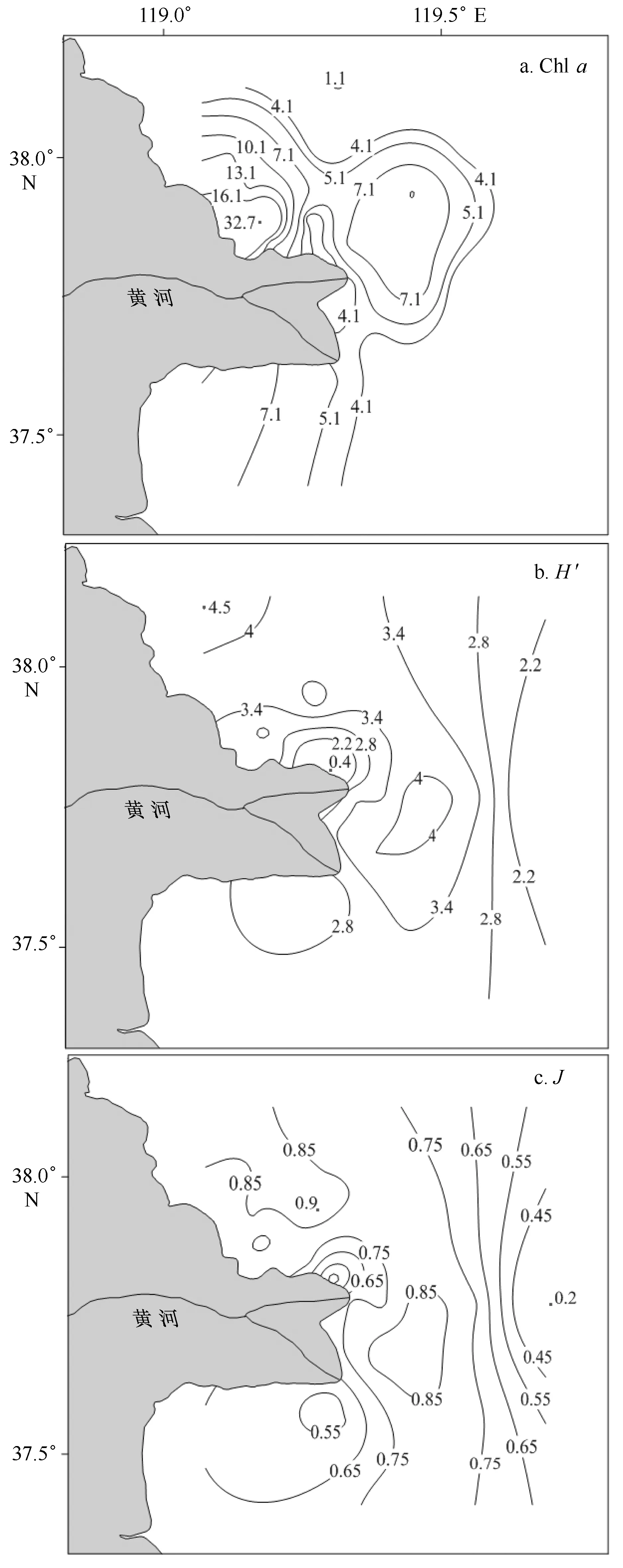
图4 浮游植物群落结构的空间分布特征Fig.4 The spatial distribution of phytoplankton assemblages during the survey
浮游植物群落结构在各断面上出现明显差异(见图5)。黄河口靠近渤海湾的A、B断面,浮游植物群落结构主要由硅藻、甲藻组成,但也出现了少量的绿藻与蓝藻;受黄河影响较大的C、D、E断面,蓝藻与绿藻的比例显著增高,硅藻、甲藻的比例明显降低;受黄河冲淡水影响较小的F断面,则完全由硅藻组成。这与吴玉霖等[43]研究发现长江径流同长江口海域浮游植物种类组成有明显关系的结果一致,盐度对浮游植物群落结构变化起到重要作用。
浮游植物细胞数量的总丰度范围2×102~1 150×102cells/L,平均值为136.9×102cells/L;A、B、C、D断面细胞数量的高值区出现在离岸较远的站位,且F2站位出现了细胞数量的最高值,这与叶绿素a浓度的分布不一致。一方面可能与浮游植物的物种组成结构有关,如F断面虽然出现较高的细胞数量,但多数为小细胞的角毛藻(Chaetocerossp.)、海链藻(Thalassiosirasp.),对叶绿素的贡献不大;另外,与样品中的泥沙含量有关,河口区域的样品中含有大量泥沙,导致计数过程中一些小细胞物种难以辨认,是造成细胞数量低的重要原因。相对而言,叶绿素a浓度在高浑浊河口可以较好地反映浮游植物生物量的变化。
3.4 浮游植物群落结构与环境因子的相关性分析
为进一步了解黄河口邻近海域浮游植物群落的空间分布规律及其决定环境因素,本研究对浮游植物群落结构与环境因子进行了PCA分析,排序图结果如图6所示。相关分析发现:盐度与营养盐是影响群落组成及其丰度的主要因素。绿藻与蓝藻细胞丰度与盐度呈极强负相关,显示出偏好低盐环境,这与离河口最远且盐度相对最高的F断面上蓝藻和绿藻细胞丰度为零的结果相对应;而甲藻丰度则与盐度呈现极强正相关,显示出偏好较高盐度的生长环境;硅藻丰度与盐度没有显著性相关,显示了其物种的多样性以及对盐度的要求比较宽泛。这一结果显示黄河调水调沙在浮游植物群落组成上,对硅藻的影响要远低于甲藻、蓝藻与绿藻。
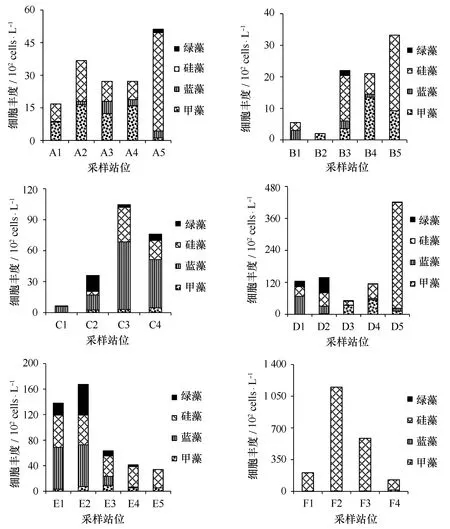
图5 浮游植物细胞丰度各断面的空间分布特征Fig.5 The spatial distribution of phytoplankton cell abundance in each transection during the survey

图6 PCA排序图:浮游植物群落结构与环境因子的相关分析Fig.6 PCA ordination plot: correlation analysis between phytoplankton assemblages and environmental factors
叶绿素a与营养盐浓度之间表现出了极强的相关性,说明了调水调沙后,营养盐大量输送对浮游植物生长的促进效应。这与在长江口的研究结果一致,例如,王云龙等[44]研究发现长江径流所携带的丰富无机氮等营养盐为浮游植物生长繁殖提供了物质基础,并通过相关性分析发现长江口海域夏季8月浮游植物总量与7月径流量呈正相关。郭玉洁和杨则禹[45]对长江口的研究也发现了径流量与浮游植物总量的正相关趋势。
通过相关分析结果,甲藻群落与硅藻群落相比,对营养盐结构的变化敏感性不高。这一现象符合甲藻的营养特征,即大部分甲藻为异养营养型,只有很少几种甲藻营严格的自养营养[46]。这可以部分解释为什么在很多氮富营养化河口、海湾,当磷酸盐与硅酸盐限制加剧时,浮游植物存在硅藻群落向甲藻群落演替的现象[47-48]。蓝藻与绿藻群落与DIP浓度表现出显著正相关,与DIN/DIP表现显著负相关;这两种类群对磷酸盐的要求相对较高[49],虽然河口大量冲淡水导致盐度降低,引起蓝藻等半咸水物种的增多,但由于受磷酸盐浓度的限制作用,难以在数量上占据主导地位。
4 结论
根据对黄河口邻近海域2013年7月调水调沙后浮游植物群落结构与环境因子的调查结果,发现:
(1)黄河水输入对河口邻近海域温度、营养盐浓度及结构形成显著影响,整体呈现由河口向离岸快速递减的分布特点,盐度呈现由河口向离岸区域逐渐递增的分布趋势。黄河口邻近海域普遍存在DIP的绝对和相对限制,DSi的限制则仅出现在局部海域。
(2)通过浮游植物与环境因子的主成分分析,发现甲藻、蓝藻与绿藻群落受盐度的影响程度明显大于硅藻群落;低盐度带来蓝藻与绿藻物种的增多,而甲藻物种的减少。在DIP与DSi限制加剧的状态下,甲藻群落对营养盐结构的敏感性低于硅藻群落。蓝藻和绿藻群落受DIP浓度的影响较大。
[1] Syvitski J P M,Vörösmarty C J,Kettner A J,et al. Impact of humans on the flux of terrestrial sediment to the global coastal ocean[J]. Science,2005,308(5720): 376-380.
[2] Walker N D,Wiseman Jr W J,Rouse Jr L J,et al. Effects of river discharge,wind stress,and slope eddies on circulation and the satellite-observed structure of the Mississippi River plume[J]. Journal of Coastal Research,2005,21(6): 1228-1244.
[3] Wang H,Yang Z,Saito Y,et al. Stepwise decreases of the Huanghe (Yellow River) sediment load (1950-2005): Impacts of climate change and human activities[J]. Global and Planetary Change,2007,57(3): 331-354.
[4] Walling D E. Human impact on land-ocean sediment transfer by the world’s rivers[J]. Geomorphology,2006,79(3): 192-216.
[5] 樊辉. 黄河口泥沙输移及三角洲的近期演变[D]. 青岛: 中国科学院海洋研究所,2005.
Fan Hui. Sediment transport in the Huanghe River mouth and the recent deltaic evolution[D]. Qingdao: Institute of Oceanology,Chinese Academy of Sciences,2005.
[6] 黄大吉,苏纪兰. 黄河三角洲岸线变迁对莱州湾流场和对虾早期栖息地的影响[J].海洋学报,2002,24(6): 104-111.
Huang Daji,Su Jilan. The effects of the Huanghe River delta on the circulation and transportation of larvae[J]. Haiyang Xuebao,2002,24(6): 104-111.
[7] Liu S M,Li L W,Zhang G L,et al. Impacts of human activities on nutrient transports in the Huanghe (Yellow River) estuary[J]. Journal of Hydrology,2012,430: 103-110.
[8] Wang Y,Liu Z,Gao H,et al. Response of salinity distribution around the Yellow River mouth to abrupt changes in river discharge[J]. Continental Shelf Research,2011,31(6): 685-694.
[9] Zhou M,Shen Z,Yu R. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River[J]. Continental Shelf Research,2008,28(12): 1483-1489.
[10] Lin C,Su J,Xu B,et al. Long-term variations of temperature and salinity of the Bohai Sea and their influence on its ecosystem[J]. Progress in Oceanography,2001,49(1): 7-19.
[11] 张焕君,李凡,丛日翔,等. 黄河口海域无脊椎动物群落结构及其变化[J]. 中国水产科学,2014,21(4): 800-809.
Zhang Huanjun,Li Fan,Cong Rixiang,et al. Community structure of invertebrate and its change in Huanghe (Yellow River) estuary[J]. Journal of Fishery Sciences of China,2014,21(4): 800-809.
[12] 吕振波,李凡,曲业兵,等. 2010年夏季黄河口及邻近海域鱼类群落多样性[J]. 渔业科学进展,2013,34(2): 10-18.
Lyu Zhenbo,Li Fan,Qu Yebing,et al. Fish community diversity in the Huanghe estuary and its adjacent area in summer,2010[J]. Progress in Fishery Sciences,2013,34(2):10-18.
[13] 刘晓彤,刘光兴. 2009年夏季黄河口及其邻近水域网采浮游植物的群落结构[J]. 海洋学报,2012,34(1): 153-162.
Liu Xiaotong,Liu Guangxing. Net-phytoplankton community structure of the Huanghe estuary and its adjacent area in the summer of 2009[J]. Haiyang Xuebao,2012,34(1): 153-162.
[14] Lorenzen C J. Determination of chlorophyll and pheopigments: spectrophotometric equations[J]. Limnol. Oceanogr,1967,12(2): 343-346.
[15] 孙军,刘东艳,钱树本. 一种海洋浮游植物定量研究分析方法——Utermöhl方法的介绍及其改进[J]. 黄渤海海洋,2002,20(2): 105-112.
Sun Jun,Liu Dongyan,Qian Shuben. A quantative research and analysis method for marine phytoplankton: An introduction to Utermöhl method and its modification[J]. Journal of Oceanography of Huanghai & Bohai Seas,2002,20(2): 105-112.
[16] 郭玉洁,钱树本. 中国海藻志[M]. 北京: 科学出版社,2003.
Guo Yujie,Qian Shuben. Flora Algarummari-Narum Sinicarum[M]. Beijing: Science Press,2003.
[17] 李伟新,朱仲嘉,刘凤贤. 海藻学概论[M]. 上海: 科学技术出版社,1982.
Li Weixin,Zhu Zhongjia,Liu Fengxian. The Introduction to Algae[M]. Shanghai: Science and Technology Press,1982.
[18] 钱树本,刘东艳,孙军. 海藻学[M]. 青岛: 中国海洋大学出版社,2005.
Qian Shuben,Liu Dongyan,Sun Jun. Marine Phycology[M]. Qingdao:China Ocean University Press,2005.
[19] 郭皓. 中国近海赤潮生物图谱[M]. 北京: 海洋出版社,2004.
Guo Hao. Illustrations of Planktons Responsible for the Blooms in Chinese Coastal Waters[M]. Beijing:China Ocean Press,2004.
[20] 杨世民,董树刚. 中国海域常见浮游硅藻图谱[M]. 青岛: 中国海洋大学出版社,2006.
Yang Shimin,Dong Shugang. Illustrations of Common Planktonic Diatoms in China Seas Waters[M]. Qingdao: China Ocean University Press,2006.
[21] 杨世民,李瑞香. 中国海域甲藻扫描电镜图谱[M]. 北京: 海洋出版社,2014.
Yang Shimin,Li Ruixiang. Atlas of Dinoflagellates in the China’s Seas[M]. Beijing: China Ocean Press,2014.
[22] 杨世民,李瑞香,董树刚. 中国海域甲藻 Ⅰ[M]. 北京: 海洋出版社,2014.
Yang Shimin,Li Ruixiang,Dong Shugang. Dinoflagellates in the China’s Seas I[M]. Beijing: China Ocean Press,2014.
[23] Shannon C E,Weaver W. The mathematical theory of communication (Urbana,IL)[J].Champaign: University of Illinois Press,1949.
[24] Pielou E C. An introduction to mathematical ecology[M]. Hoboken: John Wiley & Sons Inc,1969.
[25] 廖巍. 黄河口水质污染现状及评价指标体系的构建[D]. 青岛: 中国海洋大学,2013.
Liao Wei. Water pollution situation and establishment of evalution index system in the Yellow River estuary[D]. Qingdao: Ocean University of China,2013.
[26] 廖巍,张龙军,陈洪涛,等. 2001—2011年黄河口营养盐变化及入海通量估算[J]. 中国海洋大学学报,2013,43(1): 81-86.
Liao Wei,Zhang Longjun,Chen Hongtao,et al. Nutrients variations and fluxes estimation in the Yellow River estuary from 2001 to 2011[J]. Periodical of Ocean University of China,2013,43(1): 81-86.
[27] 黄大吉,苏纪兰,张立人. 渤海冬夏季环流的数值研究[J]. 空气动力学学报,1998,16(1): 115-121.
Huang Daji,Su Jilan,Zhang Liren. Numerical study of the winter and summer circulation in the Bohai Sea[J]. Acta Aerodynamica Sinica,1998,16(1): 115-121.
[28] 沈志良. 渤海湾及其东部水域的水化学要素[J]. 海洋科学集刊,1999,41: 51-59.
Shen Zhiliang. Hydrochemical elements in Bohai Bay and its eastern part waters[J]. Studia Marina Sinica,1999,41: 51-59.
[29] 殷鹏,刘志媛,张龙军. 2009年春季黄河口附近海域营养盐状况评价[J]. 海洋湖沼通报,2011,2: 120-130.
Yin Peng,Liu Zhiyuan,Zhang Longjun. Evalution of the nutrition status for the adjacent waters of the Yellow River estuary[J]. Transactions of Oceanology and Limnology,2011,2: 120-130.
[30] 孙晓庆,董树刚,汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产,2008,4(1): 1-9.
Sun Xiaoqing,Dong Shugang,Tang Zhihong. Influences of nutrients and illuminance on phytoplankton community structure[J]. South China Fisheries Science,2008,4(1): 1-9.
[31] 曲克明,陈碧鹃,袁有宪,等. 氮磷营养盐影响海水浮游硅藻种群组成的初步研究[J]. 应用生态学报,2000,11(3): 445-448.
Qu Keming,Chen Bijuan,Yuan Youxian,et al. A preliminary study on influence of N and P on population constituent of planktonic diatoms in seawater[J]. Chinese Journal of Applied Ecology,2000,11(3): 445-448.
[32] Ning X,Lin C,Su J,et al. Long-term environmental changes and the responses of the ecosystems in the Bohai Sea during 1960—1996[J]. Deep-Sea Research Part Ⅱ: Topical Studies in Oceanography,2010,57(11): 1079-1091.
[33] 于志刚,米铁柱,谢宝东,等. 二十年来渤海生态环境参数的演化和相互关系[J]. 海洋环境科学,2000,19(1): 15-19.
Yu Zhigang,Mi Tiezhu,Xie Baodong,et al. Changes of the environmental parameters and their relationship in recent twenty years in the Bohai Sea[J]. Marine Environmental Science,2000,19(1): 15-19.
[34] 阚文静,张秋丰,石海明,等. 近年来渤海湾营养盐变化趋势研究[J]. 海洋环境科学,2010,29(2): 238-241.
Kan Wenjing,Zhang Qiufeng,Shi Haiming,et al. Study on variation trend of nutrient salts in Bohai Bay[J]. Marine Environmental Science,2010,29(2): 238-241.
[35] 李桂菊,马玉兰,李伟,等. 春季渤海湾营养盐分布及潜在性富营养化评价[J]. 天津科技大学学报,2012,27(5): 22-27.
Li Guiju,Ma Yulan,Li Wei,et al. Distribution of inorganic nutrients and potential eutrophication assessment in Bohai Bay in spring[J]. Journal of Tianjin University of Science &Technology,2012,27(5): 22-27.
[36] Nelson D M,Brzezinski M A. Kinetics of silicic acid uptake by natural diatom assemblages in two Gulf Stream warm-core rings[J]. Marine Ecology Progress Series,Oldendorf,1990,62(3): 283-292.
[37] Justi D,Rabalais N N,Turner R E,et al. Changes in nutrient structure of river-dominated coastal waters: stoichiometric nutrient balance and its consequences[J]. Estuarine,Coastal and Shelf Science,1995,40(3): 339-356.
[38] 栾青杉,孙军. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系[J]. 生态学报,2010,30(18): 4967-4975.
Luan Qingshan,Sun Jun. Feature of phytoplankton assemblages in Yangtze River estuary and its relationship with environmental factors in summer 2005[J]. Acta Ecologica Sinica,2010,30(18): 4967-4975.
[39] 赵冉,白洁,孙军,等. 2006年夏季长江口及其邻近水域浮游植物群集[J]. 海洋湖沼通报,2009,2: 88-96.
Zhao Ran,Bai Jie,Sun Jun,et al. Phytoplankton assemblages in Yangtze River estuary and its adjacent water in summer,2006[J]. Transactions of Oceanology and Limnology,2009,2: 88-96.
[40] 田家怡. 黄河三角洲附近海域浮游植物多样性[J]. 海洋环境科学,2000,19(2): 38-42.
Tian Jiayi. Diversity of phytoplankton in the sea near Yellow River delta[J]. Marine Environmental Science,2000,19(2): 38-42.
[41] 冷宇,赵升,刘霜,等. 黄河口海域夏季浮游植物的分布特征[J]. 水生态学杂志,2013,34(6): 41-46.
Leng Yu,Zhao Sheng,Liu Shuang,et al. Distribution characteristics of phytoplankton in the estuary of Yellow River during summer[J]. Journal of Hydroecology,2013,34(6): 41-46.
[42] 朱兰部,张法高. 1989年夏季黄河口及其附近海域某些自然环境特征及水型分布[J]. 海洋科学集刊,1994,35: 23-31.
Zhu Lanbu,Zhang Fagao. Some natural environment characteristics and distribution of seawater types in the Yellow River estuary and its nearby coastal in summer 1989[J]. Studia Marina Sinica,1994,35: 23-31.
[43] 吴玉霖,傅月娜,张永山,等. 长江口海域浮游植物分布及其与径流的关系[J]. 海洋与湖沼,2004,35(3): 246-251.
Wu Yulin,Fu Yuena,Zhang Yongshan,et al. Phytoplankton distribution and its relation to the runoff in the Changjiang (Yangzte) estuary[J]. Oceanologia et Limnologia Sinica,2004,35(3): 246-251.
[44] 王云龙,袁骐,沈新强. 长江口及邻近海域夏季浮游植物分布现状与变化趋势[J]. 海洋环境科学,2008,27(2): 169-172.
Wang Yunlong,Yuan Qi,Shen Xinqiang. Distribution status and change tendency of phytoplankton during summer in Changjiang estuary and adjacent waters[J]. Marine Environmental Science,2008,27(2): 169-172.
[45] 郭玉洁,杨则禹. 长江口浮游植物的数量变动及生态分析[J]. 海洋科学集刊,1992,33: 167-189.
GuoYujie,Yang Zeyu. Quantity variation and ecological analysis of phytoplankton in the Changjiang estuary[J]. Studia Marina Sinica,1992,33: 167-189.
[46] 孙军,郭术津. 甲藻的异养营养型[J]. 生态学报,2011,31(20): 6270-6286.
Sun Jun,Guo Shujin. Dinoflagellate heterotrophy[J]. Acta Ecologica Sinica,2011,31(20): 6270-6286.
[47] Yang Dongfang,Gao Zhenhui,Sun Peiyan,et al. Mechanism of nutrient silicon and water temperature influences on phytoplankton growth[J]. 2006.
[48] 林峰竹,吴玉霖,于海成,等. 2004年长江口浮游植物群落结构特征分析[J]. 海洋与湖沼,2008,39(4): 401-410.
Lin Fengzhu,Wu Yulin,Yu Haicheng,et al. Phytoplankton community structure in the Changjiang estuary and its adjacent waters in 2004[J]. Oceanologia et Limnologia Sinica,2008,39(4): 401-410.
[49] 许海,朱广伟,秦伯强,等. 氮磷比对水华蓝藻优势形成的影响[J]. 中国环境科学,2011,31(10): 1676-1683.
Xu Hai,Zhu Guangwei,Qin Boqiang,et al. Influence of nitrogen-phosphorus ratio on dominance of bloom-forming cyanobacteria (Microcystisaeruginosa)[J]. China Environmental Science,2011,31(10):1676-1683.
附录:
Response of phytoplankton assemblages to the water-sediment regulation in the adjacent sea of the Yellow River mouth
Su Zhijuan1,2,Wang Yujue2,Dong Zhijun2,Zhang Yiwen1,Liu Dongyan2,Wang Yanxia3
(1.HebeiKeyLaboratoryofEnvironmentalEvolutionandEcologicalConstruction,CollegeofResourcesandEnvironmentScience,HebeiNormalUniversity,Shijiazhuang050024,China;2.KeyLaboratoryofCoastalEnvironmentalProcessesandEcologicalRemediation,YantaiInstituteofCoastalZoneResearch,ChineseAcademyofSciences,Yantai264003,China;3.InstituteofGeographicalScience,HebeiAcademyofScience,Shijiazhuang050021,China)
An ecological survey was carried out in July 2013 in the Yellow River adjacent sea after the 16th water-sediment regulation,with an aim of understanding the response of phytoplankton assemblages to changed environment. The result showed that temperature and nutrient concentrations decreased gradually from the mouth of river to offshore waters,corresponding to the increased freshwater loading,and salinity increased gradually from the mouth of river to offshore waters. Nutrient concentrations at the sites near to the side of Bohai Bay displayed higher values than the Laizhou Bay. The analysis on nutrient structure indicated that DIP was absolute and relative limitation,while DSi limitation only appeared in a few sites. 100 species of phytoplankton in 60 genera of the five classes were identified from the samples,and major phytoplankton components were diatoms and dinoflagellates. In addition,a few brackish species in Cyanophyceae,Chrysophyceae,and Chlorophyceae were also identified from the river mouth,indicating the impact of salinity. Chlorophyllaconcentrations and nutrient concentrations presented a significantly spatial distribution from the river mouth to offshore waters,with higher values at the sites near to the side of the Bohai Bay than the Laizhou Bay. Phytoplankton assemblages were in response to the variations of spatial environmental factors. The biomass proportions of Cyanophyceae and Chlorophyceae increased distinctly in the transections of C,D and E,indicating a strong influence of the diluted water,while diatoms and dinoflagellates in other transections with less impact from the diluted water. The results of Principal Components Analysis between phytoplankton assemblages and environmental factors indicated that DSi,DIP and salinity were key factors related to the spatial variation of phytoplankton assemblages.
Yellow River mouth; phytoplankton; water-sediment regulation; salinity; nutrients

附表1 2013年夏季黄河口邻近海域浮游植物种类名录

续附表1

续附表1
10.3969/j.issn.0253-4193.2015.04.006
2014-09-18;
2014-12-26。
中国科学院战略先导研究项目(XDA11020405);中国科学院重点部署项目(KZZD-EW-14);河北省科学院科技计划项目(13128);中国科学院院地合作项目——“河北省海岸带受损生态系统评估及修复技术研究”。
苏芝娟(1989—),女,河北省威县人,主要研究方向为数字国土与区域规划。E-mail:suzhijuan1989@163.com
*通信作者:刘东艳,研究员,主要从事海洋藻类生态学研究。E-mail:dyliu@yic.ac.cn
Q948.8
A
0253-4193(2015)04-0062-14
苏芝娟,王玉珏,董志军,等. 调水调沙后黄河口邻近海域浮游植物群落响应特征[J]. 海洋学报,2015,37(4):62—75,
Su Zhijuan,Wang Yujue,Dong Zhijun,et al. Response of phytoplankton assemblages to the water-sediment regulation in the adjacent sea of the Yellow River mouth[J]. Haiyang Xuebao,2015,37(4):62—75,doi:10.3969/j.issn.0253-4193.2015.04.006
猜你喜欢
吉林大学学报(地球科学版)(2024年3期)2024-06-03 13:22:07
黄河黄土黄种人(2023年6期)2023-08-18 19:06:49
音乐天地(音乐创作版)(2022年3期)2022-06-14 09:33:20
海洋科学(2022年12期)2022-03-01 07:25:40
金桥(2021年11期)2021-11-20 06:37:18
金桥(2021年11期)2021-11-20 06:37:08
学与玩(2019年8期)2019-10-29 03:32:16
资源节约与环保(2019年10期)2019-01-21 00:22:48
中国酿造(2014年9期)2014-03-11 20:21:07
华北水利水电大学学报(自然科学版)(2014年5期)2014-02-27 13:29:55
