
2011—2020年调水调沙前后黄河口海域浮游动物年间变化及影响因子
2022-03-01 07:25:40王秀霞高彦洁张孝民李少文杨艳艳徐炳庆王育红
海洋科学 2022年12期
王秀霞, 高彦洁, 左 明, 张孝民, 李少文, 杨艳艳, 徐炳庆, 李 凡, 王育红
2011—2020年调水调沙前后黄河口海域浮游动物年间变化及影响因子
王秀霞1, 高彦洁2, 左 明3, 张孝民1, 李少文1, 杨艳艳1, 徐炳庆1, 李 凡1, 王育红1
(1. 山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室, 山东 烟台 264006; 2. 鲁东大学 滨海生态高等研究院, 山东 烟台 264025; 3. 东营市海洋发展研究院, 山东 东营 257091)
依据2011—2020年(2016年中断)黄河调水调沙工程实施前、后黄河口海域18个站位野外调查数据, 研究浮游动物种类组成和丰度变化及优势种更替, 以查明黄河口海域浮游动物的年间变动规律及调水调沙工程的影响。结果显示: 共记录浮游动物82种类(包括浮游幼体16类), 其中刺胞动物种类数最多, 共记录26种, 其次是桡足类出现23种。浮游动物种类组成的年间变化差异显著(<0.05), 2017年(未进行调水调沙)最低; 调水调沙前、后分别记录浮游动物69种和75种, 调水调沙前后种类数呈正相关(=0.684,=0.042); 调水调沙前后优势种更替率较高; 浮游动物丰度在2011—2015年较高, 2017—2020年相对较低, 调水调沙后浮游动物丰度年间变化达极显著水平(<0.01), 而调水调沙前后没有显著差异; 浮游动物丰度水平分布总体呈现从黄河口近岸到离岸递减趋势, 密集区位于黄河口北部近岸海域, 调水调沙后密集中心较调水调沙前向离岸海域略有推移。冗余分析表明, 对黄河口浮游动物年间变化影响较大的因子是叶绿素浓度、溶解氧浓度和水温。研究表明: 2011—2020年调水调沙期间黄河口海域浮游动物丰度年间变动大于调水调沙前后变动, 调水调沙实施对浮游动物影响持续时间较短。
浮游动物; 年际变化; 调水调沙; 黄河口
河口是陆海交互的集中地带, 由于受人类活动、径流、潮流和风浪等因素共同影响, 河口海域生态环境复杂多变且敏感脆弱[1]。黄河口海域位于渤海湾和莱州湾之间, 属于典型的河口生态系统, 其饵料丰富, 是鱼类重要的繁育场[2]。黄土高原水土流失严重使得黄河沙多水少, 水沙不平衡, 下游因泥沙淤积而成为“地上悬河”, 为解决这一问题, 黄河水利委员会自2002年起利用大型水库和调度手段实施黄河调水调沙(the water and sediment discharge regulation, WSDR), 旨在通过人造洪峰冲刷河槽, 将大量水沙集中输送入海, 达到协调水沙平衡效果[3-4]。在调水调沙的影响下, 冲淡水、营养盐、泥沙等短期内急剧增加对黄河口海域生态环境及生物资源产生较大影响[5-8]。
在河口生态系统中, 浮游动物是重要的生境变化指示生物[9]。其种类组成受径流和潮流的交汇作用较为复杂[10-11], 群落结构特征亦与河口的潮汐[12]、水文[13]、营养盐[14]等环境因子以及径流[15]密切相关。因此, 研究浮游动物群落特征变化有助于全面了解河口生态系统的结构和功能变化。
关于黄河口海域浮游动物群落结构特征的研究已有较多报道[16-20]。已有的研究认为调水调沙可加速黄河中下游浮游植物的物种演替, 浮游动物生物量亦随之增加[21], 但在调水调沙实施过程中其数量和生物量变化存在波动, 有呈先升高后降低趋势[22]。本研究基于2011—2020年间调水调沙期间黄河口海域长期监测数据, 研究此季浮游动物种类组成、时空分布和优势种的年际变动, 比较调水调沙对浮游动物组成的影响, 为评估和预测黄河口海域生态系统的现状和演变趋势提供基础, 为科学实施调水调沙提供参考。
1 材料与方法
1.1 调查海区
调水调沙一般从6月下旬开始运行, 历时约20 d, 于2011—2020 年(2016年除外)调水调沙前和调水调沙后(表1)分别对黄河口海域18个调查站进行监测调查, 调查范围为37°40′N~38°09′N、119°00′E~ 119°40′E(图1)。2017年未实施调水调沙工程, 在相应时间进行调查作为对比参考。依据与黄河入海口的距离布设4个断面, 其中断面Ⅰ(包括B1、C1、D1站)距入海口5 km, 断面Ⅱ(包括A2、B2、C2、D2、E2站)距入海口10 km, 断面Ⅲ(包括A3、B3、C3、D3、E3站)距入海口20 km, 断面Ⅳ(包括A4、B4、C4、D4、E4站)距入海口40 km。

表1 2011—2020年黄河口调查时间
注: 2017*表示2017年未实施调水调沙工程, 同期在该海域进行了2次调查
1.2 样品采集及分析
采用浅海Ⅰ型浮游生物网(网孔径为505 μm, 网长度为145 cm, 网口面积为0.2 m2)从底至表垂直拖网采集浮游动物样品。样品采集后用5%的甲醛海水溶液固定保存, 带回实验室分析。水温(temperature, TEM)、盐度(salinity, SAL)和溶解氧(dissolved oxygen,DO)数据利用美国Orion 520M-01A便携式YSI水质分析仪现场测定, 叶绿素(chlorophyll, Chl)浓度采用分光光度法测定, 取1 000 mL海水经GF/F滤膜抽滤, 滤膜冷冻保存, 带回实验室测定。浮游动物样品在Stemi 508体视显微镜下鉴定、计数, 计数结果转化为丰度, 用单位海水中个体数(ind./m3)表示。采样和室内分析方法按GB/T127636—2007《海洋调查规范第6部分: 海洋生物调查》进行[23]。

图1 黄河口调查站位
黄河径流量(discharge, DIS)和输沙量(sediment load, SED)数据来自水利部黄河水利委员会发布的黄河泥沙公报(http://www.yrcc.gov.cn/zwzc/gzgb/gb/ nsgb/), 厄尔尼诺指数(Oceanic Nino Index, ONI)数据来自美国国家海洋与大气管理局的气候预测中心(https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php)。
1.3 数据分析
优势度():=(n/)f, (1)
其中,为浮游动物总个体数,n为第种个体数,f为第种出现的频率, 选取优势度≥0.02的物种作为优势种,、分别为两个相邻时期优势种数,为两个相邻时期共同优势种数。
通过R语言中vegan软件包的冗余分析(redundancy analysis, RDA)方法分析浮游动物丰度与环境因子的关系, 平面分布图运用Surfer 14(Golden Software, USA)绘制, 多元方差分析(MANOVA)检验利用IBM SPSS Statistics 19 完成。
2 结果
2.1 种类组成年际变化
2011—2020年调水调沙前后黄河口海域共记录浮游动物82种(包括浮游幼体16种), 其中刺胞动物种类数最多, 共记录26种, 占总种类数的31.71%, 其次是桡足类出现23种, 占28.05%。调水调沙前调查共记录浮游动物69种, 分属于原生动物门、刺胞动物门、节肢动物门、毛颚动物门和脊索动物门等, 以节肢动物门种类数最多, 共计录29种, 其中桡足类21种类, 占总种类数的30.43%, 刺胞动物门和浮游幼体次之, 各出现21种和14类, 分别占总种类数的30.43%和20.29%。调水调沙后调查浮游动物种类数较调水调沙前增多, 共记录75种, 种类组成与调水调沙前调查结果相似, 以节肢动物门种类数最多, 共计33种, 占总种类数的44%, 其中桡足类22种, 占29.33%; 其次是刺胞动物门, 出现22种(占29.33%), 浮游幼体16种(占21.33%)。
如图2显示, 浮游动物种类数在调水调沙前与调水调沙后的整体变化趋势相似, 差异不显著(MANOVA,1, 308=0.007,=0.933), 但其年间波动较为明显, 特别是在2017年, 浮游动物种类数<20, 减少幅度最大, 年间变化差异显著(8, 308=24.502,< 0.01)。

图2 2011—2020年调水调沙前后浮游动物丰度和种类数年际变化
2.2 优势种年间变化
黄河口海域浮游动物优势种(表2)中, 强壮滨箭虫()、中华哲水蚤()、小拟哲水蚤()、拟长腹剑水蚤()、双壳类幼体和长尾类糠虾幼体优势度平均值较高, 强壮滨箭虫作为优势种的出现频率最高, 除2012年调水调沙后调查外, 它是所有调查季的优势种。
调水调沙前后优势种更替率较高(图3), 在2012年和2017年均为81.82%, 优势种类丰度在调水调沙前后存在显著差异(=0.04)。细颈和平水母仅在2017年7月调查时成为优势种(=0.12), 出现频率为100%, 在C3站位丰度最高, 达10.42 ind./m3。图4显示, 浮游动物主要优势种类丰度的年间变化梯度较站位间更明显, 相对高丰度值出现时间不同, 但其多分布在黄河入海口近岸海域的断面Ⅰ和断面Ⅱ站位。

图3 2011—2020年调水调沙前后浮游动物优势种更替率年间变化
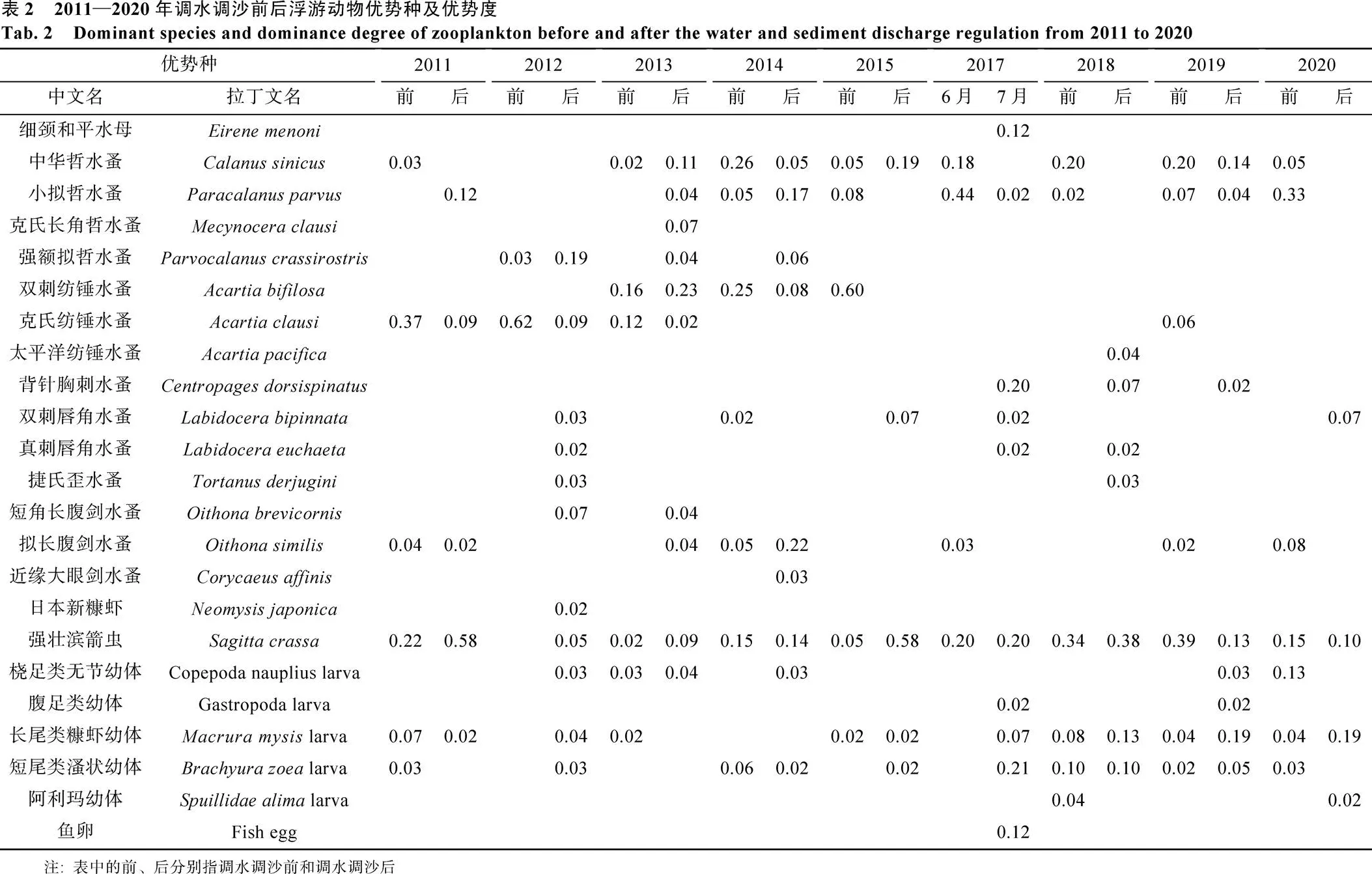
图4 主要优势种类丰度(ind./m3)的时空分布
Fig. 4 Temporospatial abundance (ind./m3) distributions of main dominant species
注: 图中的前、后分别指调水调沙前和调水调沙后, 下同
2.3 丰度时空变动
比较而言, 无论是调水调沙前还是调水调沙后, 2011—2015年海域的浮游动物丰度较高, 2017—2020年相对较低(图2)。多元方差分析显示, 浮游动物丰度年间差异显著(8, 308=11.255,<0.01), 但在调水调沙前的年间差异不显著(>0.05), 而在调水调沙后年间差异极显著(<0.001)。浮游动物在调水调沙后的丰度大多低于调水调沙前, 仅在2011年和2014年有所不同, 但浮游动物丰度在调水调沙前、后差异性并不显著。
不同年间, 浮游动物丰度分布略有差异, 但总体分布趋势相似, 即高丰度值多分布在紧邻黄河入海口处。对比各年调水调沙前、后的浮游动物丰度分布, 在调水调沙前它主要呈现从黄河口近岸断面Ⅰ到离岸断面Ⅳ递减趋势, 密集区位于黄河口北部近岸海域; 在调水调沙后, 浮游动物丰度密集区从近岸海域向离岸海域推移, 在断面Ⅱ和断面Ⅲ相对密集。2020年与其他年份不同, 调水调沙前浮游动物丰度呈现从西北到东南逐渐升高的趋势, 密集中心位于E4站位, 而在调水调沙后密集区向近岸移动至D2站位附近(图5)。
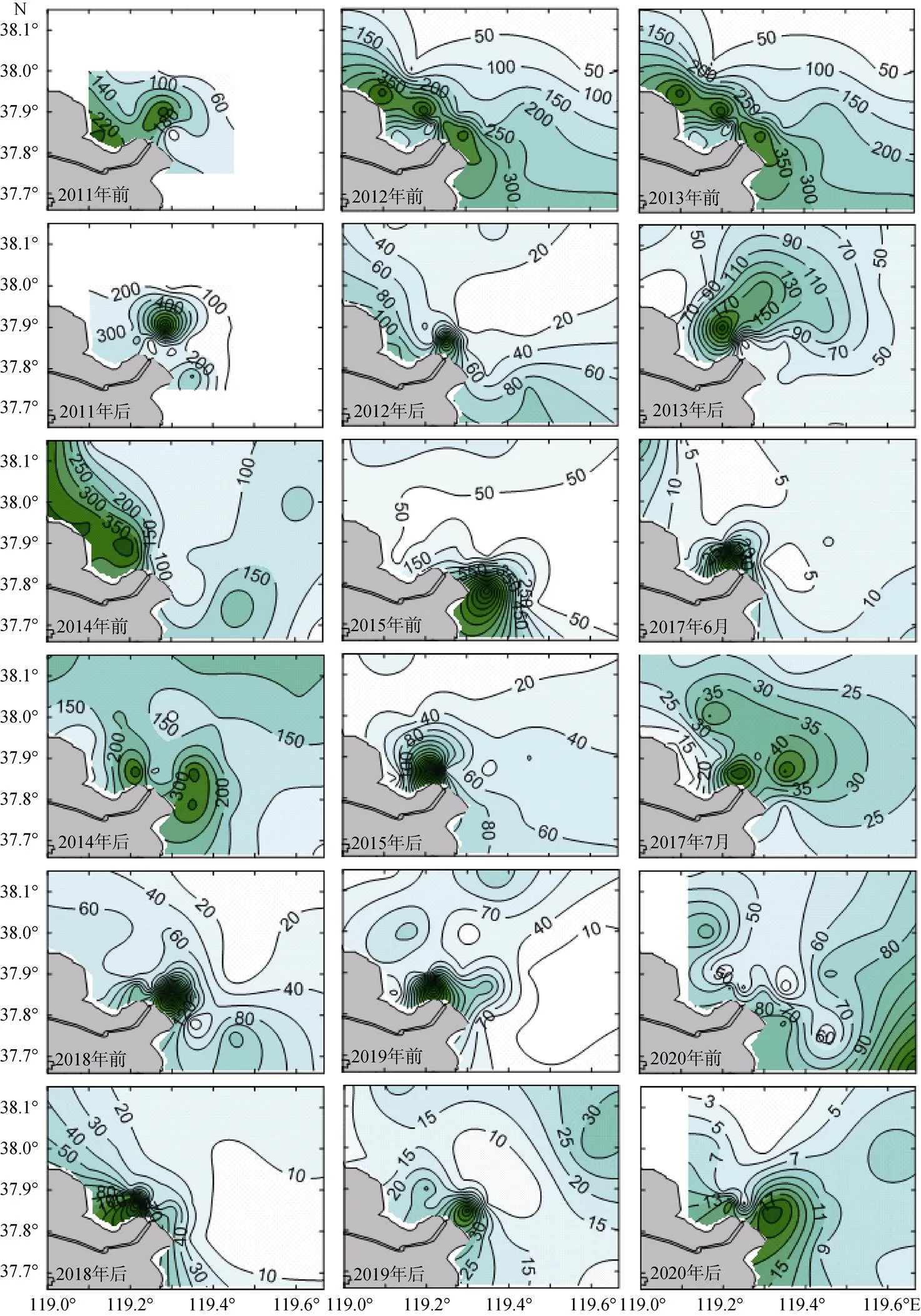
图5 2011—2020年浮游动物丰度(ind./m3)水平分布
2.4 年间变动的影响因子分析
2011—2020年调查期间, 黄河口海域水温介于15.74~30.05 ℃, 调水调沙后水温明显高于调水调沙前(图6); 多元方差分析(MANOVA)显示, 调查海域水温年间变化和调水调沙前后变化均达极显著水平(8, 308=30.713,<0.01;8, 308=506.348,<0.01)。盐度在2011—2017年较为稳定, 平均盐度为28.8, 少数河口站位盐度较低, 2018年盐度降低, 且分布较为离散, 调水调沙前和调水调沙后调查平均盐度分别为24.5和23.1; 盐度年间差异极显著(8, 308=9.103,< 0.01), 而在调水调沙前后差异不显著(8, 308=0.151,=0.698)。调查期间黄河口海域溶解氧总体波动幅度较小, 变化范围为4.58~11.73 mg/L, 年间差异和调水调沙前后差异均不显著。叶绿素浓度变化范围为0.11~13.60 μg/L, 年间变化显著(8, 290=13.067,< 0.01), 尤其以2015年调水调沙前调查和2013年调水调沙后调查与同期其他年份差异较大; 调水调沙前后没有明显差异。
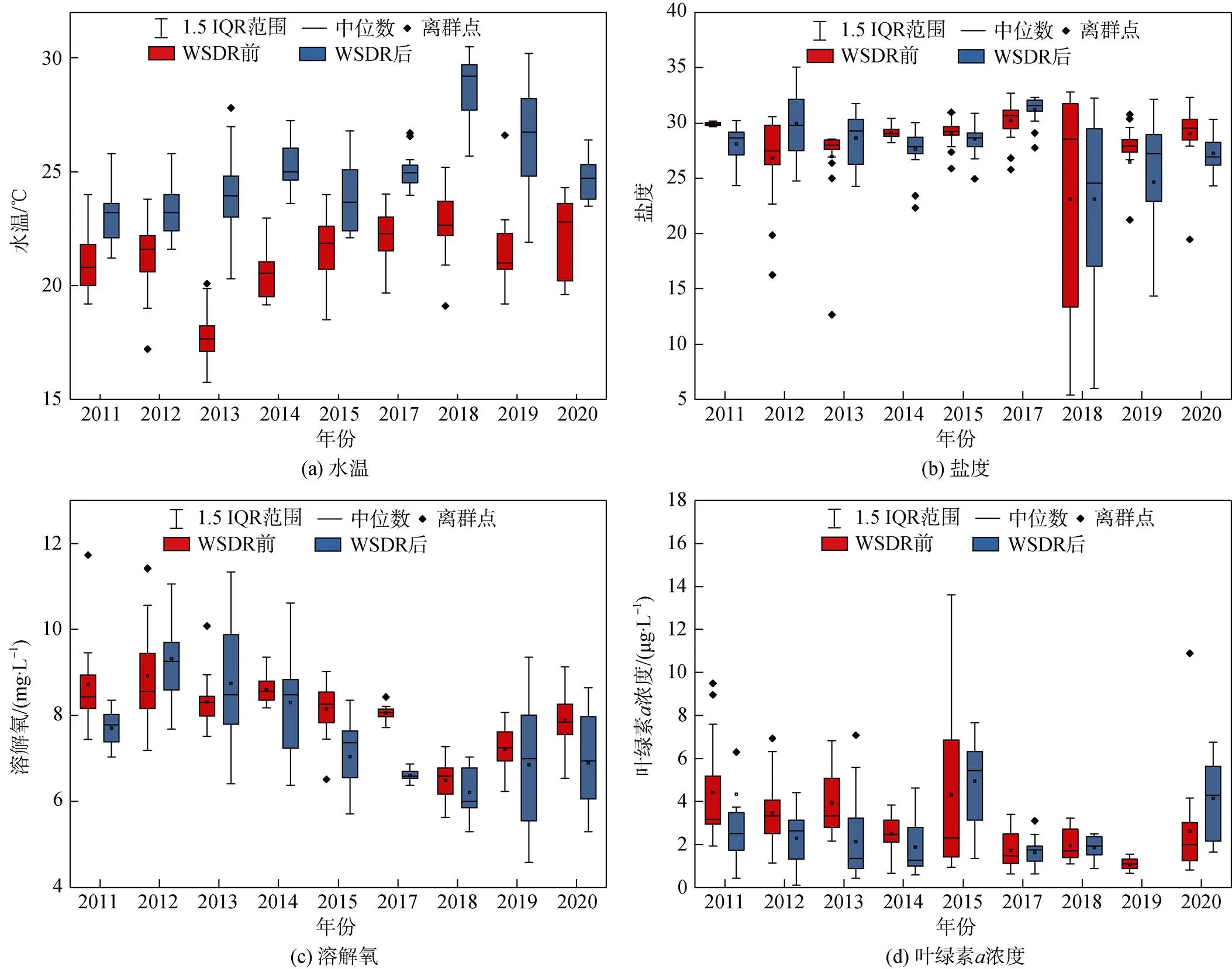
图6 调水调沙前后黄河口海域环境因子年间变化
黄河径流量和输沙量变化趋势一致, 年间变化均达显著水平(<0.05), 2018—2020年相比其他年份较高(图7)。调水调沙期间径流量和输沙量年内变动幅度较大, 多在7月迅速达到峰值, 2017年未进行调水调沙, 径流量和输沙量年内变动较小。图5所示, 厄尔尼诺指数连续5个月滑动平均值超过+0.5 ℃, 则认为发生一次厄尔尼诺事件, 若连续5个月滑动平均值低于−0.5 ℃, 则认为发生一次拉尼娜事件, 由此判断2010、2011、2020年发生拉尼娜事件, 2015年发生一次厄尔尼诺事件。

图7 黄河利津站径流量、输沙量及厄尔尼诺指数年际变动
冗余分析结果显示(图8, 表3), 第一轴RDA1的解释比例是42.91%, 第二轴RDA2的解释比例是9.80%, 根据环境因子和排序轴的相关系数, 对浮游动物群落年间变化影响较大的环境因子是底层叶绿素浓度、水温和溶解氧(图8a), 第一轴RDA1 相关系数分别为0.267、−0.201 和−0.066, 第二轴RDA2 相关系数为−0.088、0.180 和−0.255。水温和叶绿素浓度对2014年WSDR前、2015年WSDR前、2019年WSDR前和2018年WSDR后影响明显, 溶解氧对2012年和2013年影响明显。径流量、输沙量、盐度和厄尔尼诺指数对浮游动物群落年际变化也有一定的影响, 径流量和输沙量对调水调沙前后影响较大, 盐度和厄尔尼诺指数对2014年WSDR前、2015年WSDR前、2019年WSDR前后有一定影响。对中华哲水蚤和小拟哲水蚤丰度变动影响较大的是水温、径流量和输沙量, 对拟长腹剑水蚤丰度变动影响较大的是叶绿素浓度和径流量, 强壮滨箭虫丰度分布与盐度、溶解氧和叶绿素浓度相关性较大, 而长尾类糠虾幼体和短尾类溞状幼体分布主要受水温、径流量和输沙量影响。
3 讨论
2011—2020年调水调沙后调查记录的浮游动物种类数略高于调水调沙前调查, 均以刺胞动物、桡足类和浮游幼体种类数较高, 刺胞动物(水母类)种类数占比有所升高, 调查期间黄河口海域共记录浮游动物82种类, 刺胞动物种类数(26种)超过桡足类(23种), 这与近年来夏季黄河口海域浮游动物调查结果一致。2007年夏季黄河口海域浮游动物调查共记录浮游动物72个种类(包括浮游幼体20类), 其中水母类17种, 占总种类组成的23.6%, 超过桡足类(16种, 占22.2%)[11]。2009年夏季调查黄河口海域浮游动物共记录65种类, 其中桡足类16种, 水母类15种, 分别占25%和23%[17]。董志军等[20]对2012—2013年黄河口邻近海域中小型浮游动物调查结果表明, 桡足类种类最多(21种), 占总种数的30%; 其次是水螅水母类(15种), 占总种数的21.4%。刘镇盛[24]、杜明敏等[25]研究结果显示, 夏季渤海海域浮游动物群落结构发生了变化, 水母类种类数超过桡足类, 在浮游动物中所占比例最高, 在本研究中, 水母类出现的种类数高于桡足类, 但桡足类丰度较高, 在总丰度中的优势地位未改变。

图8 浮游动物丰度的影响因子分析
注: TEM表示水温; SAL表示盐度; DO表示溶解氧; Chl表示叶绿素浓度; DIS表示径流量; SED表示输沙量; ONI表示厄尔尼诺指数; 前、后分别代表WSDR前和WSDR后

表3 环境因子与排序轴之间的相关系数
与以往报道相似, 黄河口海域浮游动物优势种主要是近岸低盐种和广温广盐种[11, 17, 19-20]。调水调沙前调查海域浮游动物的优势种主要有中华哲水蚤、双刺纺锤水蚤、克氏纺锤水蚤、小拟哲水蚤和强壮滨箭虫等, 随着黄河调水调沙的实施, 径流量和输沙量骤增, 短期内大量的淡水、泥沙等急剧增加使黄河口海域生态环境发生改变[5], 在调水调沙后水温升高(图6), 优势种出现了太平洋纺锤水蚤、背针胸刺水蚤、双刺唇角水蚤和真刺唇角水蚤等近岸低盐暖温种。在2017年7月调查中水母类细颈和平水母成为优势种, 该种在渤海历史调查中夏季数量较多, 偶尔在水母类中成为优势种[17, 26-27], 但成为浮游动物优势种尚未见于报道。细颈和平水母属于近岸暖水种, 偏喜略高水温, 在7月份以前主要分布于渤海三大湾内和沿岸水域, 7月份后明显向中央海区扩展[26], 2011年调查在莱州湾内出现于6—10月, 逐月从湾底西侧逐渐向整个湾区扩展, 7月仅黄河口近岸3个站位出现[27]。在本研究中, 2017—2019年细颈和平水母均有出现, 在2018年和2019年调水调沙后调查分别有3个和4个站位出现, 但2017年7月调查出现频率达100%, 平均丰度为3.73 ind./m3。2016—2017年调水调沙工程中断, 入海水沙量显著减少, 黄河口海域海洋动力相对增强, 等深线向陆地蚀退[28], 2017年黄河径流量与其他调查年份相比较低(图5), 盐度较高(图4), 有利于细颈和平水母生长, 而浮游动物种类数和丰度整体较低(图2), 导致细颈和平水母成为优势种。
调水调沙前调查海域浮游动物密集区主要位于黄河入海口口门北侧近岸低盐、高营养盐的浅水区水域, 远岸深水区丰度相对较低, 呈现从近岸到远岸降低的趋势。主要是因为数量较大的双刺纺锤水蚤、克氏纺锤水蚤、强壮滨箭虫等为近岸低盐种, 集中分布于近岸海域, 远岸深水区盐度相对较高, 以小拟哲水蚤、拟长腹剑水蚤等广温广盐种为主, 且丰度较低。随着调水调沙的实施, 大量淡水集中流入黄河口近岸海域, 同时将全年60%以上的泥沙和50%以上的营养物质携带入海[29-30], 黄河入海径流量增大, 径流作用强于潮流, 冲淡水沿河道主轴线向口门东北方向前进, 受地球自转和海流共同影响形成明显的顺时针环流和低盐中心[31]。调水调沙后冲淡水范围退缩, 但依旧呈舌状向外海延伸[5], 浮游动物密集中心向外海略有推移, 浮游动物以近岸低盐种和广温广盐种为主, 高盐外海种亦有出现, 表明黄河口海域浮游动物受黄河冲淡水和黄海海流共同影响, 调水调沙实施使黄河径流量短期内大量增加, 增强了黄河口海域环流, 从而将外海种带到近岸, 近岸低盐种分布范围向外海延伸。2020年调水调沙前调查海域浮游动物分布趋势与以往调查不同, 密集中心位于东南部E4站位, 以小拟哲水蚤数量最高, 这可能与该站位水温(24.3 ℃)较高有关系, 适合暖温种小拟哲水蚤生长, 调水调沙后密集区向近岸移动。
本研究RDA分析表明, 对浮游动物丰度年际变化影响较大的环境因子是底层叶绿素浓度、溶解氧和水温, 除上述因素外, 研究表明, 黄河口海域叶绿素、溶解氧、浮游植物、渔业资源等在调水调沙实施过程中短期内变动较大。营养盐和溶解氧含量随着径流量的激增而快速升高, 随着调水调沙的结束, 营养盐逐渐恢复至调水调沙前水平[30], 而溶解氧保持较高水平。浮游植物丰度则呈现先降低后升高的趋势, 调水调沙后期与调水调沙前水平接近[22]。鱼类生物量先升高后降低[6], 且渔业资源结构年际变化大于调水调沙前后变化[32]。2014年出现史上最强的厄尔尼诺事件[33], 导致2014 年、2015年全球海水温度出现异常, 并对全球海洋生物资源和全球海洋生态系统产生了重要影响[34]。本研究RDA分析亦显示, 浮游动物丰度与对厄尔尼诺、拉尼娜等大尺度气候因子有一定相关性。分析表明厄尔尼诺对2014年调水调沙前和2015 年调水调沙前浮游动物丰度呈负相关。在异常气候与黄河调水调沙双重影响下, 黄河口浮游动物产生了较明显响应。
4 小结
2011—2020年调水调沙期间黄河口海域浮游动物种类数存在显著年际差异, 浮游动物丰度在调水调沙后年际变化达极显著水平, 而浮游动物种类数和丰度在调水调沙工程实施前后均没有显著差异, 浮游动物丰度年际变化大于调水调沙前后变化。可以说明, 黄河口海域浮游动物变动主要受不同年份环境状况影响, 其次才是调水调沙的影响, 且调水调沙实施对其影响持续时间较短。
[1] 杨宇峰, 王庆, 陈菊芳, 等. 河口浮游动物生态学研究进展[J]. 生态学报, 2006, 26(2): 576-585.
YANG Yufeng, WANG Qing, CHEN Jufang, et al. Research advance in estuarine zooplankton ecology[J]. Acta Ecologica Sinica, 2006, 26(2): 576-585.
[2] 杨尧尧, 李忠义, 吴强, 等. 莱州湾渔业资源群落结构和多样性的年际变化[J]. 渔业科学进展, 2016, 37(1): 22-29.
YANG Yaoyao, LI Zhongyi, WU Qiang, et al. Interannual variations in community structure and species diversity of fishery resources in the Laizhou Bay[J]. Progress in Fishery Sciences, 2016, 37(1): 22-29.
[3] YANG H B, LI E C, ZHAO Y, et al. Effect of water- sediment regulation and its impact on coastline and suspended sediment concentration in Yellow River Estuary[J]. Water Science and Engineering, 2017, 10(4): 311-319.
[4] LI G Y, SHENG L X. Model of water-sediment regulation in Yellow River and its effect[J]. Science China Technological Sciences, 2011, 54(4): 924-930.
[5] 孙珊, 苏博, 李凡, 等. 调水调沙对黄河口及邻近海域环境状况的影响[J]. 海洋环境科学, 2019, 38(3): 399- 406.
SUN Shan, SU Bo, LI Fan, et al. Effects of water and sediment discharge regulation on environment in the Yellow River Estuary and adjacent waters[J]. Marine Environmental Science, 2019, 38(3): 399-406.
[6] 郑亮, 吕振波, 李凡, 等. 调水调沙期间黄河口海域鱼类群落结构特征[J]. 中国水产科学, 2014, 21(3): 602-610.
ZHENG Liang, LV Zhenbo, LI Fan, et al. Fish community structure in the Yellow River estuary: Effect of water and sediment discharge regulations[J]. Journal of Fishery Sciences of China, 2014, 21(3): 602-610.
[7] 苏芝娟, 王玉珏, 董志军, 等. 调水调沙后黄河口邻近海域浮游植物群落响应特征[J].海洋学报, 2015, 37(4): 62-74.
SU Zhijuan, WANG Yujue, DONG Zhijun, et al. Response of phytoplankton assemblages to the water- sediment regulation in the sdjacent sea of the Yellow River mouth[J]. Haiyang Xuebao, 2015, 37(4): 62-75.
[8] LI S W, LI F, SONG X K, et al. The influence of water- sediment regulation on macrobenthic community structures in the Huanghe River (Yellow River) Estuary during 2012-2016[J]. Acta Oceanologica Sinica, 2020, 39(10): 120-128.
[9] 杨志, 叶金清, 杨青, 等.辽河口海域浮游动物多样性及对河口环境的指示[J].海洋环境科学, 2020, 39(1): 25-30.
YANG Zhi, YE Jin-qing, YANG Qing, et al. Zooplankton diversity and its relationships with environmental factors in the Liaohe estuary[J]. Marine Environmental Science, 2020, 39(1): 25-30.
[10] 邵倩文, 刘镇盛, 章菁, 等. 长江口及邻近海域浮游动物群落结构及季节变化[J]. 生态学报, 2017, 37(2): 683-691.
SHAO Qianwen, LIU Zhensheng, ZHANG Jing, et al. Seasonal variation in zooplankton community structure in the Changjiang Estuary and its adjacent waters[J]. Acta Ecologica Sinica, 2017, 37(2): 683-691.
[11] 马静, 陈洪举, 刘光兴. 2007年夏季黄河口及其邻近水域浮游动物的群落特征[J]. 中国海洋大学学报(自然科学版), 2012, 42(5): 74-80.
MA Jing, CHEN Hongju, LIU Guangxing. Study on the zooplankton community structure in the Yellow River estuary and its adjacent waters in summer, 2007[J]. Periodical of Ocean University of China, 2012, 42(5): 74-80.
[12] MAHARA N, PAKHOMOV E A, DOSSER H V, et al. How zooplankton communities are shaped in a complex and dynamic coastal system with strong tidal influence[J]. Estuarine, Coastal and Shelf Science, 2021, 249: 107103.
[13] 王文杰, 刘光兴. 2010年秋季黄河口及其邻近海域中小型浮游动物的群落特征[J]. 海洋科学, 2013, 37(11): 9-15.
WANG Wenjie, LIU Guangxing. The characteristics of meso- and micro-zooplankton community in the Yellow River estuary and its adjacent area in autumn, 2010[J]. Marine Sciences, 2013, 37(11): 9-15.
[14] YUAN D N, CHEN L D, LUAN L L, et al. Effect of salinity on the zooplankton community in the Pearl River estuary[J]. Journal of Ocean University of China, 2020, 19(6): 1389-1398.
[15] 王晓东. 长江口浮游动物功能群的长期变化——基于时间序列和生态化学计量学的视角[D]. 青岛: 中国科学院大学(中国科学院海洋研究所), 2017.
WANG Xiaodong. Long-term change of zooplankton functional groups in the Changjiang Estuary – A view based on time-series and ecological stoichiometry[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2017.
[16] 葛汝平, 刘光兴, 陈洪举, 等. 春季黄河口两种网型网采浮游动物的群落特征[J]. 中国海洋大学学报(自然科学版), 2019, 49(4): 62-70.
GE Ruping, LIU Guangxing, CHEN Hongju, et al. Community characteristics of zooplankton sampled with two plankton nets in Yellow River Estuary in Spring[J]. Periodical of Ocean University of China, 2019, 49(4): 62-70.
[17] 李浩然, 刘光兴, 马静, 等. 夏、秋季黄河口及邻近水域浮游动物群落特征[J]. 海洋环境科学, 2018, 37(5): 631-639.
LI Haoran, LIU Guangxing, MA Jing, et al. Community characteristics of zooplankton in the Yellow River estuary and its adjacent area in summer and autumn[J]. Marine Environmental Science, 2018, 37(5): 631-639.
[18] 冷春梅, 曹振杰, 张金路, 等. 黄河口浮游生物群落结构特征及环境质量评价[J]. 海洋环境科学, 2014, 33(3): 360-365.
LENG Chunmei, CAO Zhenjie, ZHANG Jinlu, et al. Community characteristic of plankton and water quality assessment of the Yellow River Estuary[J]. Marine Environmental Science, 2014, 33(3): 360-365.
[19] 刘晓慧, 徐宾铎, 张崇良, 等. 黄河口及其邻近水域浮游动物群落结构特征及其季节变化[J]. 中国水产科学, 2017, 24(5): 922-930.
LIU Xiaohui, XU Binduo, ZHANG Chongliang, et al. Characteristics of zooplankton community structure and its seasonal variation in the Yellow River estuary and its adjacent waters[J]. Journal of Fishery Sciences of China, 2017, 24(5): 922-930.
[20] 董志军, 杨青, 孙婷婷, 等. 黄河口邻近海域浮游动物群落时空变化特征[J]. 生态学报, 2017, 37(2): 659- 667.
DONG Zhijun, YANG Qing, SUN Tingting, et al. Spatial and seasonal variability of the zooplankton community in the Yellow River Estuary’s adjacent sea[J]. Acta Ecologica Sinica, 2017, 37(2): 659-667.
[21] SONG J, HOU C Y, LIU Q, et al. Spatial and temporal variations in the plankton community because of water and sediment regulation in the lower reaches of Yellow River[J]. Journal of Cleaner Production, 2020, 261: 120972.
[22] ZHANG J J, LI F, LV Q M, et al. Impact of the water– sediment regulation scheme on the phytoplankton community in the Yellow River estuary[J]. Journal of Cleaner Production, 2021, 294: 126291.
[23] 国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763—2007, 海洋调查规范第6部分: 海洋生物调查[S]. 北京: 中国标准出版社, 2008: 1-157.
General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, China National Standardization Administration. GB/T 12763.6-2007, Specifications for oceanographic survey Part 6: Marine biological survey[S]. Beijing: China Standards Press, 2008: 1-157.
[24] 刘镇盛, 杜明敏, 章菁. 国际海洋浮游动物研究进展[J]. 海洋学报, 2013, 35(4): 1-10.
LIU Zhensheng, DU Mingmin, ZHANG Jing. International research advances in marine zooplankton[J]. Acta Oceanologica Sinica, 2013, 35(4): 1-10.
[25] 杜明敏, 刘镇盛, 王春生, 等. 中国近海浮游动物群落结构及季节变化[J]. 生态学报, 2013, 33(17): 5407- 5418.
DU Mingmin, LIU Zhensheng, WANG Chunsheng, et al.The seasonal variation and community structure of zooplankton in China sea[J]. Acta Ecologica Sinica, 2013, 33(17): 5407-5418.
[26] 马喜平, 高尚武. 渤海水母类生态的初步研究——种类组成、数量分布与季节变化[J]. 生态学报, 2000, 20(4): 533-540.
MA Xiping, GAO Shangwu. The ecology of medusae in the Bohai Sea – Species composition, quantitative distribution and seasonal variation[J]. Acta Ecologica Sinica, 2000, 20(4): 533-540.
[27] 左涛, 吴强, 王俊, 等. 莱州湾水母种类多样性及群集结构的季节变化[J]. 生态学报, 2016, 36(18): 5646- 5656.
ZUO Tao, WU Qiang, WANG Jun, et al. Annual survey of the species diversity and assemblage dynamics of medusae in Laizhou Bay, Bohai Sea[J]. Acta Ecologica Sinica, 2016, 36(18): 5646-5656.
[28] 陈俊卿, 范勇勇, 吴文娟, 等. 2016—2017年调水调沙中断后黄河口演变特征[J]. 人民黄河, 2019, 41(8): 6-9, 116.
CHEN Junqing, FAN Yongyong, WU Wenjuan, et al. Evolution characteristics of the Yellow River estuary after interruption of the Yellow River water and sediment regulation in 2016-2017[J]. Yellow River, 2019, 41(8): 6-9, 116.
[29] LIU S M, LI L W, ZHANG G L, et al. Impacts of human activities on nutrient transports in the Huanghe (Yellow River) estuary[J]. Journal of Hydrology, 2012, 430/431: 103-110.
[30] LIU S M. Response of nutrient transports to wateresediment regulation events in the Huanghe basin and its impact on the biogeochemistry of the Bohai[J]. Journal of Marine Systems, 2015, 141: 59-70.
[31] 王柯萌, 孙永福, 宋玉鹏, 等. 调水调沙对黄河三角洲切变锋及冲淡水扩散的影响[J]. 海洋科学进展, 2019, 37(4): 551-564.
WANG Kemeng, SUN Yongfu, SONG Yupeng, et al. Characteristics of shear fronts and dispersal of diluted water during the water-sediment regulation scheme in Yellow River Delta[J]. Advances in Marine Science, 2019, 37(4): 551-564.
[32] 陈宁, 杨艳艳, 魏振华, 等. 调水调沙前后黄河口渔业资源结构变化[J]. 中国水产科学, 2017, 24(5): 953- 962.
CHEN Ning, YANG Yanyan, WEI Zhenhua, et al. Composition and distribution of fishery resources before and after the water and sediment discharge regulation in the Yellow River estuary[J]. Journal of Fishery Sciences of China, 2017, 24(5): 953-962.
[33] JACOX M G, HAZEN E L, ZABA K D, et al. Impacts of the 2015-2016 El Niño on the California Current System: Early assessment and comparison to past events[J]. Geophysical Research Letters, 2016, 43(13): 7072-7080.
[34] KOGAN F, GUO W. Strong 2015-2016 El Niño and implication to global ecosystems from space data[J]. International Journal of Remote Sensing, 2017, 38(1): 161-178.
Interannual variation and influencing factors of zooplankton in the Yellow River estuary before and after water and sediment discharge regulation from 2011 to 2020
WANG Xiu-xia1, GAO Yan-jie2, ZUO Ming3, ZHANG Xiao-min1, LI Shao-wen1, YANG Yan-yan1, XU Bing-qing1, LI Fan1, WANG Yu-hong1
(1. Shandong Key Laboratory of Marine Ecological Restoration, Shandong Marine Resources and Environment Research Institute, Yantai 264006, China; 2.Institute for Advanced Study of Coastal Ecology, Ludong University, Yantai 264025, China; 3. Marine Development Research Institute, Dongying 257091, China)
The water and sediment discharge regulation (WSDR) has been implemented since 2002 to address the water and sediment imbalance in the Yellow River. To explore the response of zooplankton to the WSDR in the Yellow River estuary, we analyzed the species composition, spatiotemporal distribution, and dominant species replacement rate of zooplankton using the samples collected by type I plankton net from bottom to surface at 18 sampling stations from 2011 to 2020. The analysis was performed for two conditions: both before and after the WSDR. The results showed that 82 species of zooplankton (including 16 species of planktonic larvae) were recorded in the estuary of the Yellow River; cnidarians had the highest number with 26 species, followed by copepods with 23 species.The interannual variation of zooplankton species was the lowest in 2017 (0.05).Before and after the WSDR, 69 and 75 zooplankton species were recorded, respectively, and the replacement rate of the dominant species was high. Before the WSDR, the abundance of zooplankton was higher in 2011-2015 than in 2017-2020, with no significant difference betweenthe two time periods (before and after the WSDR).The horizontal distribution of zooplankton decreased from the coastal area to the offshore area, and the northern coastal area of the Yellow River Estuary had the highest concentration. Redundancy analysis showed that the most influential factors on zooplankton abundance in the estuary of the Yellow River were chlorophyll, dissolved oxygen, and temperature. In this study, the interannual variation of zooplankton in the estuary of the Yellow River estuary was projected to be greater than that of the WSDR from 2011 to 2020, and the impact of the WSDR on zooplankton in the estuary of Yellow River was short-lived.
zooplankton; interannual variation; the water and sediment discharge regulation; the Yellow River estuary
Apr. 2, 2022
S931
A
1000-3096(2022)12-0115-13
10.11759/hykx20220402001
2022-04-02;
2022-05-07
山东省自然科学基金重点项目(ZR2020KE050); 山东省海洋生态修复重点实验室开放课题(201902); 青岛海洋科学与技术试点国家实验室山东省专项经费(2021QNLM050103); 烟台市科技创新发展计划(2020MSGY061); 烟台市科技创新发展计划项目(2021XDHZ053)
[Key project of Shandong Provincial Natural Science Foundation, No. ZR2020KE050; Shandong Key Laboratory of Marine Ecological Restoration, No. 201902; Financially supported by the Marine S&T Fund of Shandong Province for Pilot National Laboratory for Marine Science and Technology (Qingdao), No. 2021QNLM050103; The Science and Technology Innovation Development Program of Yantai , No. 2020MSGY061; The Science and Technology Innovation Development Program of Yantai, No. 2021XDHZ053]
王秀霞(1988—), 女, 工程师, 主要从事海洋生物学研究, E-mail: xiuxia.888@163.com; 李凡(1981—),通信作者, 男, 副研究员, 主要从事渔业资源研究, E-mail: lifan811230@126.com
(本文编辑: 赵卫红)
猜你喜欢
黄河黄土黄种人(2023年6期)2023-08-18 19:06:49
黄河黄土黄种人(2023年6期)2023-08-18 19:06:49
音乐天地(音乐创作版)(2022年3期)2022-06-14 09:33:20
金桥(2021年11期)2021-11-20 06:37:18
金桥(2021年11期)2021-11-20 06:37:08
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
湖南农业(2015年7期)2015-03-20 01:48:37
华北水利水电大学学报(自然科学版)(2014年5期)2014-02-27 13:29:55
