
呼吸中间神经元及呼吸网络模型的动力学分析*
2015-05-24 03:45:05平小方刘深泉任会霞汪雷
动力学与控制学报 2015年3期
平小方 刘深泉 任会霞 汪雷
(1.华南理工大学数学系,广州 510640)(2.上海交通大学生物医学工程系,上海 200240)
引言
包含人在内的几乎所有的哺乳动物都要依靠呼吸运动来维持生命,从出生到死亡,呼吸的节律性一直持续着.由于呼吸是一种极为复杂的节律性生理活动,并且与人类息息相关,所以对呼吸节律的研究具有很重要的意义.近年来,对哺乳动物呼吸节律的研究越来越多,如 Feldman JL[1]等人提出,位于包钦格复合体与吻端VRG之间的一个区域-前包钦格复合体(PBC)可能是新生哺乳动物呼吸节律起源的关键部位,之后的研究表明成年猫[2]和成年大鼠[3]的PBC在呼吸节律的产生中也起着关键性的作用.目前关于呼吸节律的产生机制有两种学说:起搏细胞学说和神经网络学说.但由于方法学方面的限制,目前的研究更多的是集中在神经网络学说方面.在这个基础上,之后的相关研究主要集中在对特定的呼吸神经元和呼吸网络的研究上.
对呼吸神经元的研究,在大鼠及狗的PBC内有多种呼吸神经元,包括前吸气神经元、早吸气神经元、递增型吸气神经元、晚吸气神经元、吸气后神经元和递增型呼气神经元等[4].其中吸气神经元的簇发放得到了广泛的研究,并确定了簇发放的机制依赖于持续Na+电流或者细胞内Ca+浓度的变化,Toporikova和 Butera[15]建立了一个具有两种独立簇发放机制的两房室的PBC神经元模型,发现胞体中的簇发放是由持续Na+电流的失活引起的,而轴突内的簇发放依赖于由神经调节物质决定的Ca+振荡.刘义等[5]通过膜电位图和 ISI指标,分析了Pre-Bötzinger中间神经元模型中部分离子通道参数的改变对膜电位的影响,揭示了模型中蕴含着的丰富的动力学特性.Koizumi等[6]证明在试管内的大白鼠的延髓切片的Pre-B复合体中,持续Na+电流和钾电流K+对泄露电流具有支配作用,并有助于呼吸节律的产生.Butera等[7]发现当 Pre-Bötzinger中间神经元通过兴奋性氨基酸介导突触的耦合和兴奋性驱动力的突触输入控制时,神经元的发放频率得到了控制并出现同步现象.
对于呼吸网络的研究,在Smith等[8]提出的网络模型上,Rubin等[9]在一个由四个神经元和三个兴奋性驱动力构成的呼吸网络中得出:持续钠电流对三时相和二时相振荡的产生不是必要的,但是它控制着这两种状态的震荡周期[9].Rybak等[10]提出在正常呼吸期间,与呼吸相关的脑桥结构支配着髓质网络中的呼吸时相转换机制,抑制着 Pre-Bötzinger内在的起搏驱动力的振荡,并提供吸气抑制和呼气促进的条件反射.Rybak等[16]提出了两相和三相的中心呼吸模式发生器(CPRG)的网络模型,其中两相CPRG网络模型中的呼气转换机制是基于早吸气神经元和递减型呼气神经元之间相互抑制的基础上的,三相CPRG网络模型中的呼气转换机制是基于前包钦格吸气神经元的基础上的,同时不仅从细胞水平上得到了呼吸神经元的具体发放模式,而且还从网络水平上提供了呼气发放模式时相之间的转换,最后得出系统水平上可以控制呼气和吸气在不同情况下的持续时间.
在本文中,我们采用脑桥-髓质的呼吸网络模型[10],由于模型中包含了 Pre-Bötzinger中间神经元,所以,为了更好的研究呼吸网络模型中蕴含的动力学性质,我们先分析了模型中Pre-Bötzinger中间神经元放电行为的动力学特性,然后对呼吸网络模型做了一些探讨.
1 模型与方法
Hodgkin-Huxley(HH)模型是一种经典的神经元模型,也是后续一些离子通道模型产生的重要参照,本文的模型采用的也是HH型离子通道模型,下面将分别介绍 Pre-Bötzinger中间神经元模型[11-12]和我们构造的呼吸网络模型(见图1).
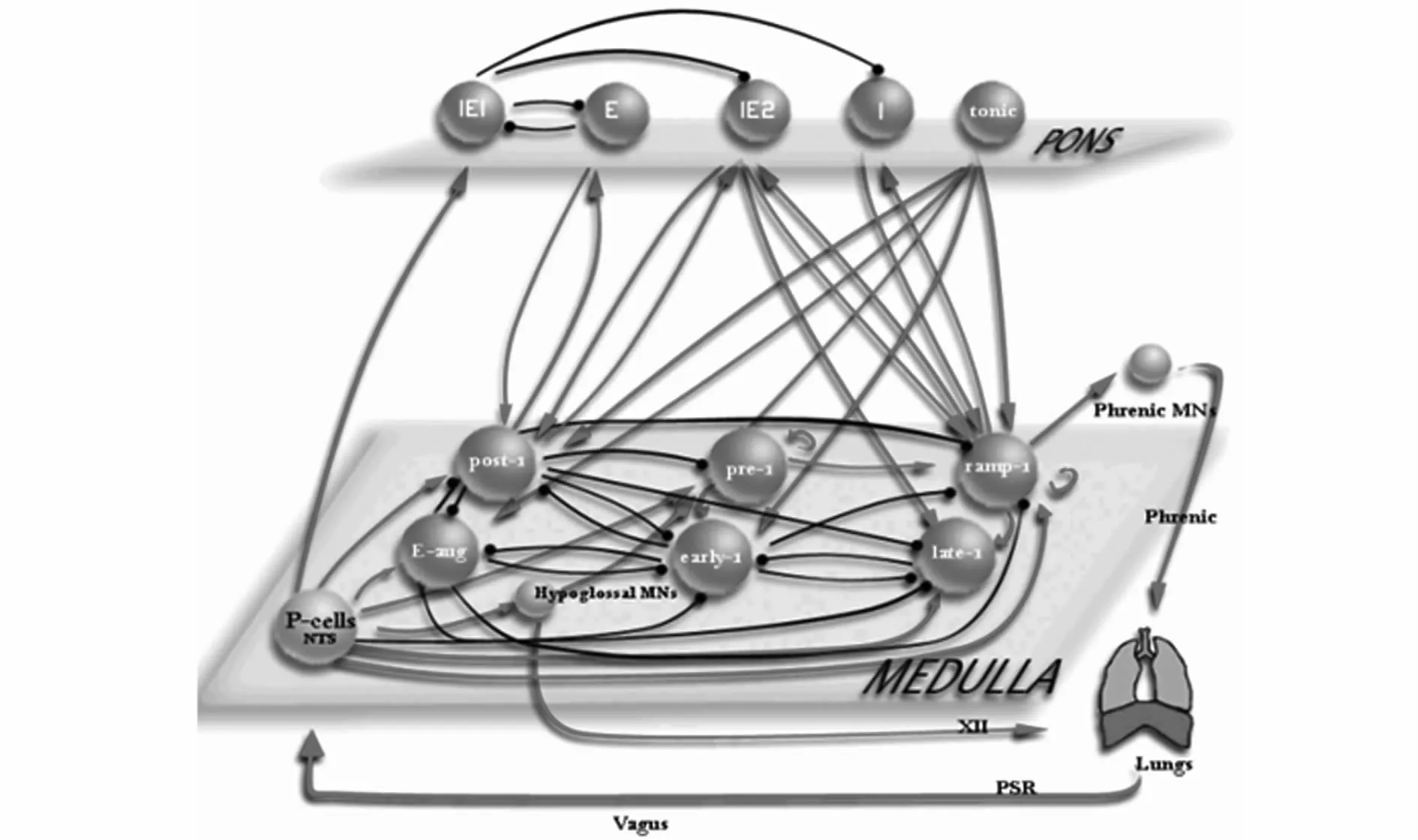
图1 脑桥—延髓呼吸网络模型Fig.1 Ponto-medullary respiratory network model
1.1 Pre-Bötzinger中间神经元模型
本节中的模型是基于单房室的Hodgkin-Huxley(HH)离子通道模型,模型的描述是由一系列微分方程构成的,具体描述如下:
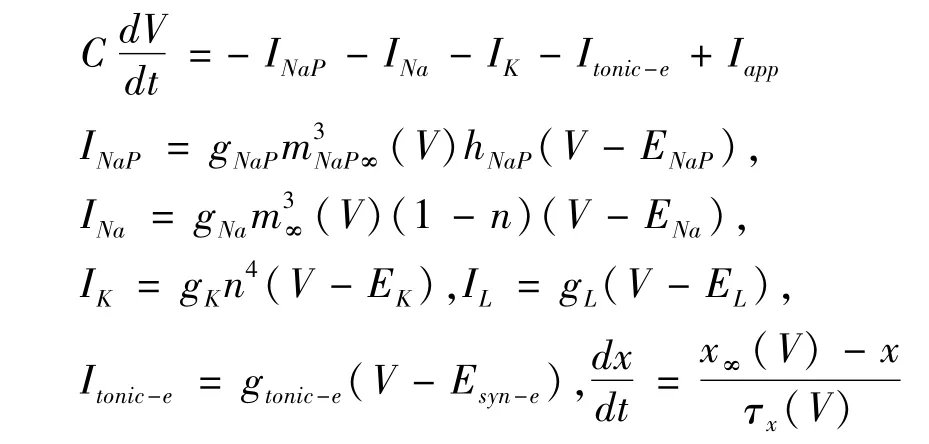
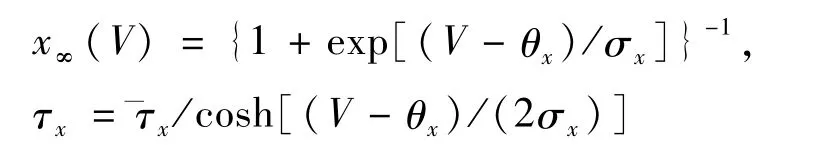
其中,C是膜电容,V是膜电位,t是时间,INaP是持续钠电流,INa是快速钠电流,IK是延迟钾电流,IL是泄露电流,Iapp是外界刺激电流,Itonic-e是受到其它神经元的兴奋性输入的突触电流.其中,门控变量x(x代表m,n,mNaP,hNaP),x∞(V)为稳态的依赖电压x(失活)的激活函数,τx(V)是依赖电压的时间常数.另外,本文参数单位规定如下:膜电容为pF,膜电位为mV,电导为nS,电流为pA,时间为ms.
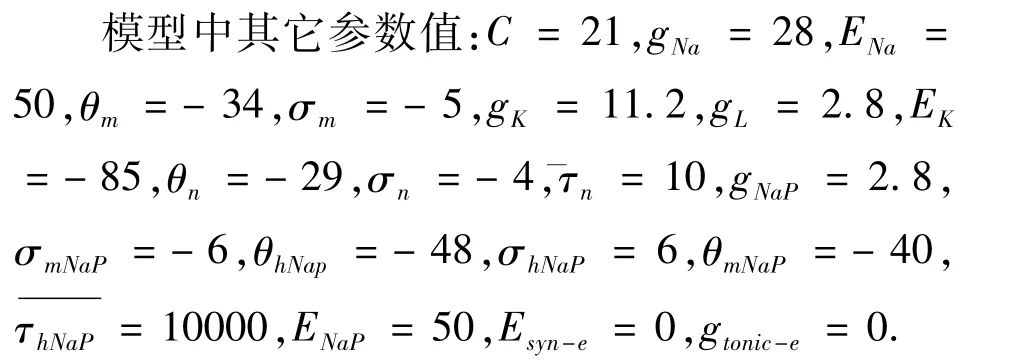
单个Pre-Bötzinger神经元模型的计算运用的是四阶龙哥库塔算法,步长为0.01ms,并通过XPP和MATLAB实现.
1.2 呼吸网络模型
本文呼吸网络模型是建立在Rybak等[10]人提出的脑桥-髓质呼吸网络模型的基础上并进行适当改进而来的,主要包含了脑桥和髓质两大部分.脑桥包含头端脑桥和尾端脑桥,其中呼气神经元组,吸气神经元组,吸-呼神经元组在头端脑桥中,驱动力(tonic)在尾端脑桥中;髓质包含头段背侧呼吸组(rVRG)、Pre-Bötzinger和 Bötzinger三部分,其中斜发放吸气神经元组和晚吸气神经元组在rVRG中,后吸气神经元组和递增型呼气神经元组在Bötzinger中,前呼气神经元组和早呼气神经元组在Pre-Bötzinger中,除此之外,呼吸网络模型中还包含了肺,孤束核中 P(Pump)细胞,膈神经运动神经元,舌下神经运动神经元.同时,呼气神经元组、吸气神经元组、吸-呼神经元组、斜发放吸气神经元组、后吸气神经元组、递增型呼气神经元组、前呼气神经元组和早呼气神经元组呼吸会受到不同大小的外界刺激,呼吸网络模型中的神经元组之间通过兴奋性突触和抑制性突触连接起来,并通过肺的输入和输出连接起来,共同构成了呼吸回路.每个呼吸神经元组中包含50个神经元,所有神经元模型均采用Hodgkin-Huxley型模型,并且假定所有神经元模型都采用Pre-Bötzinger神经元模型的形式.本文的网络模型中,圆球表示不同类型的呼吸神经元组,网络中的红色箭头和黑色圆点分别代表兴奋性和抑制性突触连接[9,11].其中:黄色代表的孤束核中P(Pump)细胞、膈神经运动神经元、舌下神经运动神经元不包含在模型的计算中.
呼吸网络模型的数值模拟实现是运用欧拉算法,步长是0.01ms,并通过 NEURON和 MATLAB软件实现.
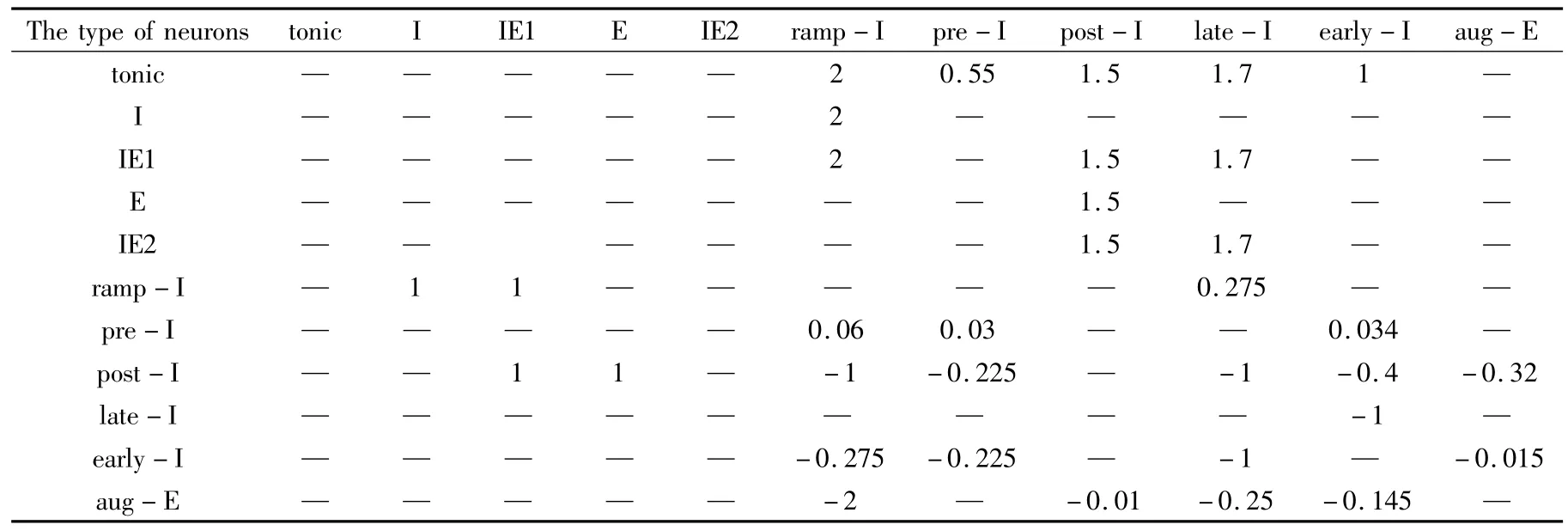
表1 呼吸网络模型中神经元组之间的突触权重Table 1 Synaptic weights between neuron groups in the respiratory network
2 模拟结果与分析
动作电位是神经元电活动的重要表现形式,其频率和波形是神经元用于将信息从一个局部传到另一个局部的编码方式,因此,动作电位在很大程度上能反应神经元的活动特性.作为动作电位的两种最基本的形式,簇放电和峰放电编码了神经元放电序列中大量的信息.本文分别通过对 Pre-Bötzinger中间神经元和呼吸网络模型中神经元膜电位变化的分析,从非线性动力学角度理解了呼吸网络中可能蕴含着的丰富的动力学现象.
2.1 Pre-Bötzinger中间神经元模型的模拟结果与分析
在早期的工作中,Rybak[14]等人对Pre-Bötzinger中间神经元出现的簇发放进行了研究,发现簇发放出现的内在原因是延迟整流器钾电流通过细胞外钾电流的增加或者持续钠电流的增加而受到抑制的结果.刘义[4]等利用膜电位变化图和膜电位的ISI等指标分析了神经元的膜电容、泄露电流的平衡电位、持续Na+的平衡电位、延迟钾K+电流的平衡电位以及它们的电导系数对神经元膜电位的影响及其存在的丰富的动力学现象.但是模型中其它的电生理参数并未讨论,为了完整全面的理解模型中可能存在着的动力学现象,下面我们将对其它参数进行讨论分析,对Pre-Bötzinger中间神经元模型的研究从神经元的动作电位入手,并结合ISI指标来共同刻画放电节律的变化,具体是通过改变快速钠的平衡电位,快速钠的电导系数和依赖电压的最大时间常量和与门控变量相关的参数对膜电位发放模式的影响来研究.从而深刻理解单个Pre-Bötzinger中间神经元模型中蕴含的动力学特性.因为Pre-Bötzinger中间神经元包含在呼吸网络模型中,所以这也为下面研究呼吸网络模型中蕴含的动力学特性打下了基础.
2.1.1 电导系数gNa对膜电位的影响
离子电流与其电导成正比,但是电导系数作为电导变化的上限在神经元动作电位的产生和转变过程中起着重要作用.在电流-电压关系中,随着膜电位趋向钠的平衡电位和驱动力的减小,钠电流将减小.但是,钠电导会随着去极化而迅速增加,并超过了驱动力的减小,从而使得钠电流增加[13].事实上,研究钠电导对膜电位的影响的文献很多[14].下面我们将通过发放图和ISI指标来研究钠电导系数对膜电位的影响.
从图2(a)中可以看出,随着快速钠平衡电位的逐渐减小,Pre-Bötzinger中间神经元的发放模式从周期性的单峰放电逐渐转化为周期性的双峰放电,接着出现了周期性的簇发放.在图(b)中,随着ENa从32减小到20,ISI序列从周期1峰发放经倍周期分岔到周期2簇放电,又经历了两次倍周期分岔,产生周期8簇放电,最后通过一个复杂放电区域进入到加周期放电序列.

图2 (a)当 gK=14时,膜电位(a)和 ISI(b)随 gNa的变化图Fig.2 (a)When gK=14,variations of membrane potentials(a)and ISIs(b)with respect to gNa
2.1.2 依赖电压的最大时间常数对膜电位的影响
电压的上升与下降用指数函数来描述.在脉冲期间的上升相由以下的函数来描述:

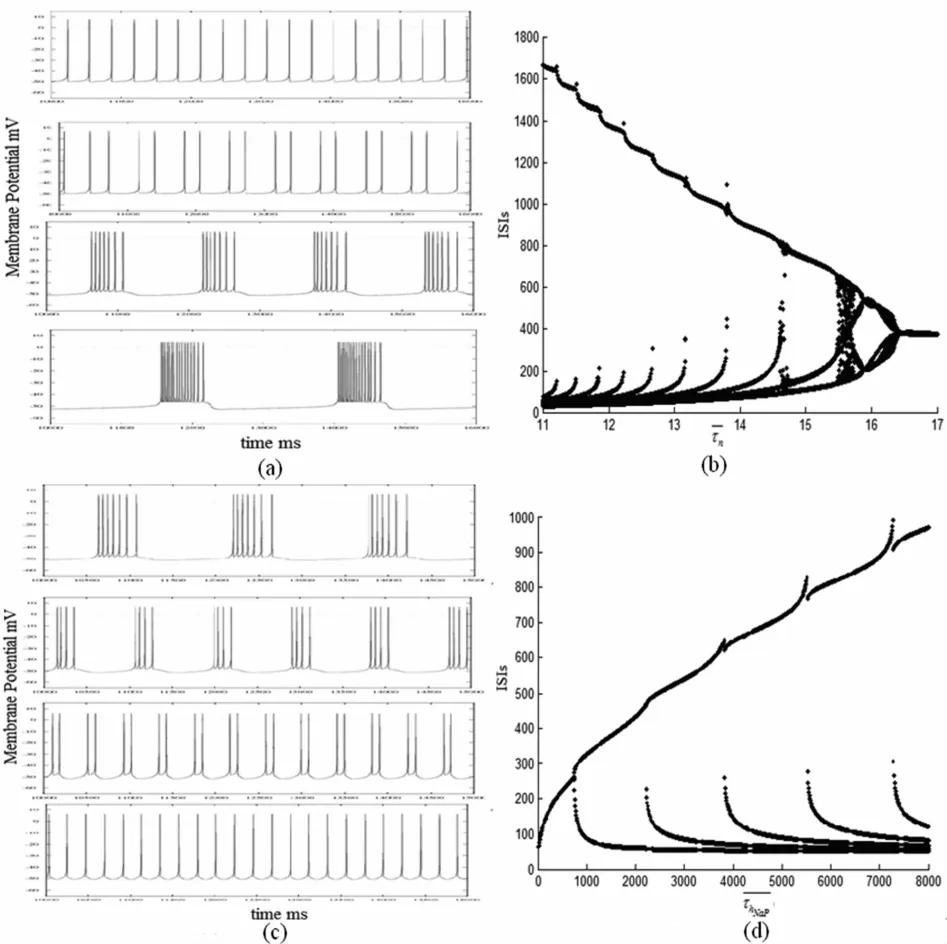
图3 膜电位(a,c)和 ISIs(b,d)随时间常数的变化图Fig.3 Variations of membrane potentials(a,c)and ISIs(b,d)with respect to
这里,t是脉冲开始后的时间.可以看出,指数时间常数τ,是由电路中的电阻和电容的乘积给出,它是电压升到其终值的1-e-1(或63%)所需要的时间.时间常数提供了一个方便的方法来测量细胞的比膜电阻.细胞膜电位达到稳态所需要的时间取决于细胞膜的时间常数.以下将对依赖电压的最大时间常数分别进行详细的动力学分析.
从图3知道,依赖电压的时间常数的改变都能引起神经元发放模式的明显变化.值得注意的是,随着参数的增加,相应的ISIs图中均出现了倍周期和加周期分岔现象,但是二者又有些不同,前者是由加周期分岔逐渐到倍周期分岔,后者是由倍周期分岔过渡到加周期分岔.这说明虽然依赖都是电压的时间常数,但是对神经元发放模式的转变扮演着不同的角色,这使得我们进一步的认识了Pre-Bötzinger中间神经元中复杂多样的放电模式以及不同模式转移中所出现的周期性分岔结构.
2.1.3 与门控变量相关的参数对膜电位的影响
在HH模型描述中,Na+离子和K+离子电导的变化由三个变量(m,h,n)决定,其中m和h的变化决定了Na+离子电流值的大小,n的变化则决定了K+离子电流值的大小,这些变量变化的快慢决定了在不同时刻离子通道电流值的不同,从而会影响神经元膜电位值的不同以及动作电位发放序列的形状.任何对变量的微小扰动,都可能会引起神经元动作电位发放模式的改变,从而影响神经元的活性.从Pre-Bötzinger中间神经元模型中知道,与门控变量相关的参数有 σm,θm,σmNaP,θmNaP,σn,θn,σhNaP和θhNaP,它们间接地影响着离子电流的大小.下面我们将通过ISI指标对这些参数进行数值分析,进而深刻理解这些参数的变化对 Pre-Bötzinger中间神经元发放模式的影响.
我们已经知道,与门控变量m相关的参数有σm,θm,θmNaP和 σmNaP,与门控变量 n相关的参数有θn和 σn,与门控变量 h相关的参数有 σhNaP和θhNaP,下面我们将分类进行分析.
从图4(a-d)可以看出,在与门控变量m相关的参数 σm,θm,θmNaP和 σmNaP的 ISI序列中,均出现了2倍周期和4倍周期分岔现象,其中a-c中还出现了加周期现象;图4(d)中更加值得注意的是,随着σmNaP从-8逐渐增加到-4,ISI序列中先出现了2倍周期分岔,接着出现了4倍周期分岔,通过一段复杂区域后,相继出现了逆4倍周期分岔和2倍周期分岔现象,这在之前的研究中是很少出现的.
从图4(e-f)知道,在与门控变量n相关的参数θn和σn的ISI序列中,也出现了很好的2倍周期分岔和4倍周期分岔以及加周期分岔现象,这其中蕴含了参数θn和σn的变动明显引起了神经元动作电位序列模式的改变和不同模式之间的转移规律.
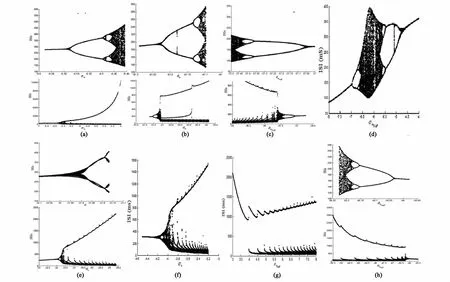
图4 ISIs随参数 σm(a),θm(b),θmNaP(c),σmNaP(d),σn,θn(e),σn(f),σhNaP(g)和 θhNaP(h)的变化图Fig.4 Variations of ISIs with respect toσm(a),θm(b),θmNaP(c),σmNaP(d),σn,θn(e),σn(f),σhNaP(g)andθhNaP(h)
从图4(g-h)知道,在与门控变量h相关的参数σhNaP和θhNaP的ISI序列中,同样有加周期现象和倍周期分岔现象.其中,图g中随着参数σhNaP的不断增加,ISI序列中仅出现了加周期现象,然而图h中,随着参数θhNaP的逐渐增加,ISI序列中不仅出现了加周期现象,还出现了倍周期分岔现象,这说明参数θhNaP的变化更能引起神经元动作电位序列丰富的变化.
通过ISI指标对上面各个参数的分析知:与门控变量相关的参数 σm,σmNaP,σhNaP,σn,θm,θn和θhNaP的改变深刻影响着膜电位,并出现了丰富的动力学特性,比如倍周期分岔和加周期等非线性动力学现象,Pre-Bötzinger中间神经元模型中蕴含的动力学现象得到更全面的诠释.那么,包含 Pre-Bötzinger中间神经元的呼吸神经网络模型中又有什么样的动力学现象呢?下面将给出具体的数值模拟分析.
2.2 呼吸网络模型的模拟结果与分析
下面是对呼吸网络模型进行的模拟和分析,主要从呼吸网络模型中各类神经元的膜电位入手,通过同步性和频率等指标来分析呼吸网络模型中蕴含的丰富的动力学性质.
2.2.1 呼吸网络模型中的发放特性
呼吸网络模型是由11类呼吸神经元和肺的相互连接共同构成的.分别给定神经元大小为0.5*(t%150<30)的方波刺激和1.5pA的直流刺激,持续时间是20s,下面是网络中各类神经元的发放图,横轴是时间,纵轴是电位.

图5 不同刺激下,网络模型中各类神经元的膜电位图Fig.5 Membrane potentials of neurons in the network under different stimuli
从图5中可以明显看出,在方波刺激和直流刺激下,呼吸网络模型中的各类神经元均出现了发放现象,但是在不同类型刺激下,一部分同类神经元的发放模式发生了变化,比如Pre-I等;另一部分同类神经元的发放模式变化不明显,比如tonic等,这反映了呼吸神经元对刺激类型的敏感性.同时,在同种刺激下,不同类型神经元的反应不同,表明不同类型的呼吸神经元对外界传入刺激的敏感性.
2.2.2 呼吸网络模型中的同步现象
Butera[7]等人对两个耦合的同样的起搏神经元进行模拟时,发现随着兴奋性驱动力的增加,同步簇发放的频率也随之增加;然而,增加两个神经元之间的兴奋性耦合强度,却降低了同步簇发放的频率.本文将从直观的发放图入手,进一步分析呼吸网络模型中的同步现象.
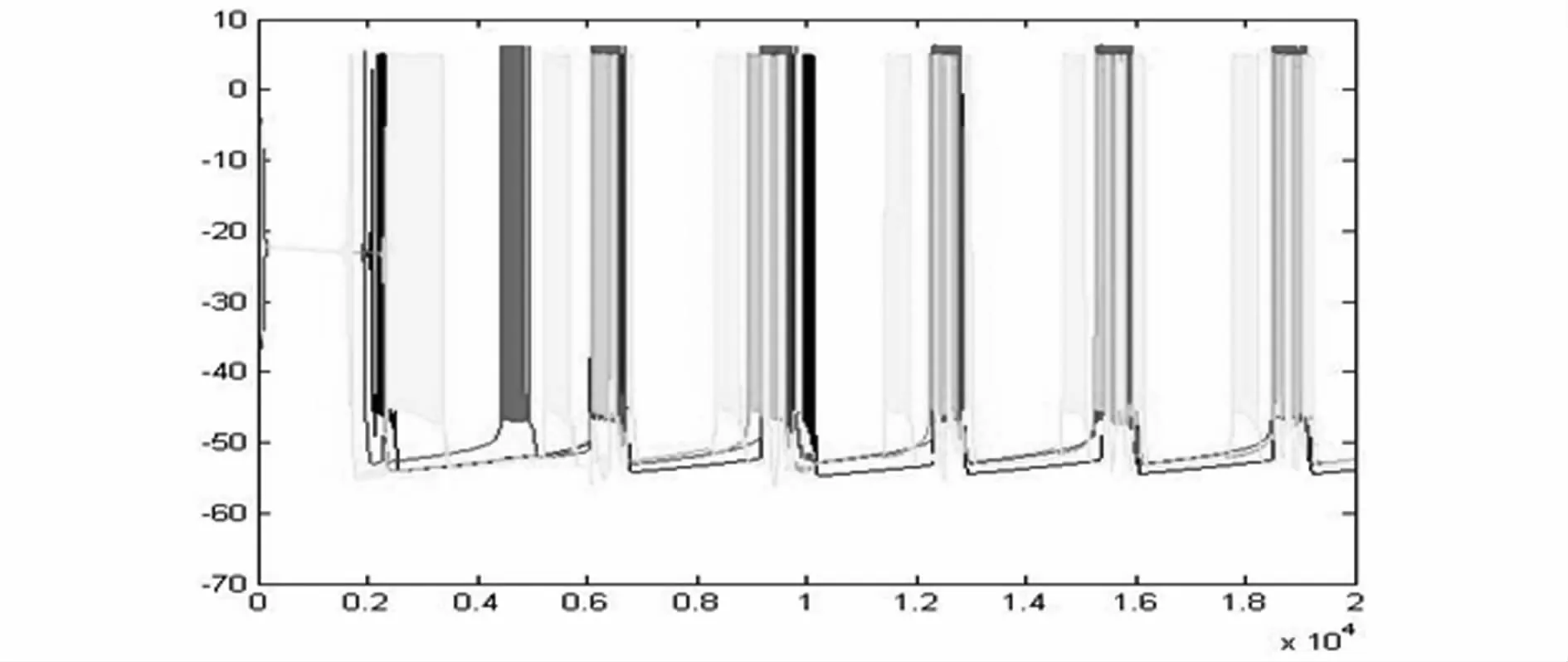
图6 不同类型神经元的膜电位图Fig.6 Membrane potentials of different neurons

图7 不同突触权重下,呼吸神经元网络的同步图Fig.7 Synchronized activities of respiratory network under different synaptic weights
本文以前包钦格吸气神经元,斜发放吸气神经元,早吸气神经元和递增型呼气神经元的发放图为例来说明呼吸网络模型中存在着同步现象和不同步现象.从图6中知道,递增型呼气神经元(绿色)和前包钦格吸气神经元(红色)相对于早吸气神经元(黄色)有一定的延迟,斜发放吸气神经元(黑色)相对于递增型呼吸神经元有明显的延迟现象,这里的延迟就代表了某一时刻的不同步状态;但是从图6中还可以清楚看出,在某一时刻四种颜色有重合的部分,这里的重合就代表了某一时刻的同步状态.这说明呼吸网络模型在不同的时刻各类神经元可能同步,也可能不同步,为了清楚全面的了解呼吸网络模型中的同步和不同步现象,我们得到了图7中的结果,其中横坐标是神经元的个数,纵坐标是时间,单位是ms.
从图7可以看出,在每一幅图中,同一时刻(即在纵轴任选一时间,若对应于横轴的一行是同一种颜色则表示这些神经元同步,若不是同一种颜色则表示不同步),颜色不发生变化就代表此刻呼吸神经元同步,颜色发生变化则代表呼吸神经元不同步,然而同步分三种情况,即均是红色(膜电位小于-45),绿色(膜电位大于3)和蓝色(膜电位介于-45与3),不同步分四种情况,即为红蓝,红绿,蓝绿和红蓝绿,从图7(d)中可以清楚看出.突触权重值发生变化时,呼吸网络模型中的同步现象不同.(a)(权重为0.05),(b)(权重为0.08),(c)(权重为0.11)图中在不同的时刻均有同步状态,也有不同步的状态,但是不同步的状态又有所区别.从这组模型结果中我们同样可以得出,在呼吸网络中,同步和不同步是同时存在的,它们之间相互制约.不同步状态的存在会避免网络过于同步化,造成信息的冗余.而同步状态的存在又会使得信息能够得到有效地编码和传递.此外,呼吸网络中同步和不同步这两种状态之间的转变也是今后需要进一步研究和讨论的重点.前面对单个神经元在不同电生理参数下表现出来的周期性的发放模式,也为进一步考察网络中同步和不同步现象提供了参考.
2.2.3 呼吸网络模型中呼吸频率的刻画
我们知道呼吸是一种节律性的生理活动,而这种节律性可以通过呼吸频率来刻画.下面我们将利用呼吸频率来描述呼吸网络模型中蕴含的节律性.我们得到图8,其中(a)图中,坐标轴从上到下,从左到右分别表示各类神经元在6500 ms内的发放个数,78组发放图的数据和各类神经元(其中1-12分别代表斜发放吸气神经元,兴奋性驱动力神经元,前包钦格吸气神经元,递增型呼气神经元,呼气神经元,吸气神经元,吸-呼神经元(第一时相),吸-呼神经元(第二时相),早吸气神经元,肺,后吸气神经元,晚吸气神经元);(b)图是(a)图的俯视图.其中,方波的刺激范围:0.5*(6500%150<30)-4.35*(6500%150<30).
从图8(a)中知道,当方波刺激大小不断变化时,不同类型的呼吸神经元的呼吸频率各不相同,其中在6500ms内,肺的发放个数相对最少,但频率相对最大;而早吸气神经元的发放个数相对最多,但频率相对最小.从图8(b)中可以清楚的看到,同种类型呼吸神经元的频率变化不明显,但是从图8(a)中可以看到有一下凸凹部分(图中箭头处),这说明同种类型呼吸神经元的频率的确是有变化的.进而表明方波刺激对呼吸频率的影响的确是存在的,但是影响不是十分明显.
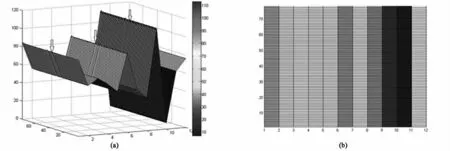
图8 方波刺激下,网络中各类神经元的频率变化图Fig.8 Variations of frequency of different neurons under square-wave stimulus
3 结论
哺乳动物的呼吸运动是由低位脑干中的呼吸神经元构成的神经网络产生的有节律的神经活动,神经元电信号有局部分级电位和动作电位两种,研究呼吸神经元动作电位的放电特性对研究电信号在神经网络中的传导有重要意义,进而对呼吸节律的研究具有重要的启示.本文中我们分别对Pre-Bötzinger中间神经元模型和呼吸网络模型进行了非线性动力学分析,得到以下几点结论:
首先,从Pre-Bötzinger中间神经元模型中得到的结论有:
(1)随着电导系数 gNa的增加,Pre-Bötzinger中间神经元膜电位的放电模式从峰放电过渡到簇放电,这个变化过程通过ISI序列可以很好的得到体现,ISI序列出现了加周期和倍周期分岔的动力学现象.
(2)随着依赖电压的最大时间常数的增加,Pre-Bötzinger中间神经元的发放序列中均出现了峰放电和簇放电模式,同时ISI序列中也均出现了加周期和倍周期分岔现象.
(3)分别对参数 σm,σmNaP,σhNaP,σn,θm,θn,θhNaP和θmNaP进行了动力学分析,通过ISI指标来刻画参数变化引起的膜电位的变化,以上各个参数的变化都使得ISI序列中出现了分岔现象,但是不同参数对应的ISI序列不同.参数σn的ISI序列中出现了2倍周期分岔和加周期现象;参数σhNaP的ISI序列中仅出现了加周期分岔现象;参数θm,σm和θn的ISI序列中均出现了2倍周期分岔和4倍周期分岔现象以及加周期分岔现象;参数 θhNaP和θmNaP的ISI序列中均出现了逆4倍周期现象和逆2倍周期分岔现象以及加周期分岔现象;参数σhNaP的ISI序列中既出现了2倍周期分岔和4倍周期分岔现象,同时也出现了逆4倍周期现象和逆2倍周期分岔现象.这充分说明了与门控变量相关的参数的变化对神经元膜电位变化的深刻影响并蕴含着丰富的动力学性质.
从以上结论可知,Pre-Bötzinger中间神经元模型中蕴含了丰富的动力学性质,这为研究呼吸网络模型中的动力学特性奠定了基础.
其次,从呼吸网络模型中得到的结论有:
(1)呼吸网络模型中的呼吸神经元在受到方波刺激和直流刺激时,一部分呼吸神经元膜电位的放电模式发生了明显的变化,另一部分呼吸神经元膜电位的放电模式变化不明显;这说明呼吸网络模型中的呼吸神经元对不同刺激的敏感性.同时,在同种刺激条件下,不同类型的呼吸神经元的放电模式也呈现出明显的不同,这说明不同类型的呼吸神经元对刺激的敏感性.
(2)以前包钦格吸气神经元,斜发放吸气神经元,早吸气神经元和递增型呼气神经元的发放图为例说明呼吸网络模型中存在同步现象和不同步现象.通过呼吸网络模型中的同步图,进一步的看清了呼吸网络模型中的同步现象和不同步现象,并且随着突触权重的变化,同步现象和不同步现象有明显的变化.
(3)当方波刺激变化时,呼吸网络模型中的各类型呼吸神经元的频率互不相同,但是同类型呼吸神经元的频率变化不是很明显,这提示了方波刺激对呼吸网络模型中的呼吸频率的影响虽然有,但是不很明显.
综上,从动力学的角度分析了Pre-Bötzinger中间神经元模型和脑桥-髓质呼吸网络模型中蕴含的丰富的动力学性质,这对以后对呼吸节律的研究提供了一定帮助,但是文章依旧存在着很多的不足,比如:呼吸节律产生的详细机制等.随着科技的不断进步和发展,研究方法和手段将不断丰富,相信通过研究者不懈的努力,对呼吸节律产生机制的研究将会不断地得到完善.
1 Feldman JL,and Del Negro C A.Looking for inspiration:new perspectives on respiratory rhythm.Nature Review Neuroscience,2006,7(3):232~242
2 Schwarzacher SW,Smith JC,Richter DW.Pre-Bötzinger complex in the cat.The Journal of Neurophysiology,1995,73(4):1452~1461
3 Sun Q J,Goodchild A K,Chalmers JP,Pilowsky P M.The Pre-Bötzinger complex and phase-spanning neurons in the adult rat.Brain Research,1998,809(2):204~213
4 张承武,郑煜.前包钦格复合体:产生呼吸节律的关键部位.生理科学进展,2002,33(2):179~181(Zhang CW,Zheng Y.Pre-Bötzinger complex:the key part of respiratory rhythm generation.Progress in Physiological Sciences,2002,33(2):179~181(in Chinese))
5 刘义,刘深泉.Pre-Bötzinger中间神经元模型的动力学分析.动力学与控制学报,2011,9(3):257~262(Liu Y,Liu SQ.Dynamic analysis of interneuron model in Pre-Bötzinger complex.Journal of Dynamics and Control,2011,9(3):257~262(in Chinese))
6 Koizumi H,Smith JC.Persistent Na+and K+-Dominated leak currents contribute to respiratory rhythm generation in the Pre-Bötzinger complex in vitro.The Journal of Neuroscience,2008,28(7):1773~1785
7 Butera R J Jr,Rinzel J,Smith JC.Models of respiratory rhythm generation in the Pre-Bötzinger complex.II.populations of coupled pacemaker neurons.The Journal of Neurophysiology,1999,82(1):398~415
8 Smith JC,Abdala A P,Koizumi H,Rybak IA,Paton J F.Spatial and functional architecture of the mammalian brainstem respiratory network:a hierarchy of three oscillatorymechanisms.The Journal of Neurophysiology,2007,98(6):3370~3387
9 Rubin JE,Shevtsova N A,Ermentrout G B,Smith JC,Rybak IA.Multiple rhythmic states in amodel of the respiratory central pattern generator.The Journal of Neurophysiol,2009,101(4):2146~2165
10 Rybak IA,Shevtsova N A,Paton JF,Dick TE,St-John W M,Mörschel M,Dutschmann M.Modeling the pontomedullary respiratory network.Respiratory Physiology&Neurobiology,2004,143(2-3):307~319
11 Butera R JJr,Rinzel J,Smith JC.Models of respiratory rhythm generation in the Pre-Bötzinger complex.I.Bursting pacemaker neurons.The Journal of Neurophysiology,1999,82(1):382~397
12 Rybak I A,Paton J F,Schwaber J S.Modeling neural mechanisms for genesis of respiratory rhythm and pattern.I.Models of respiratory neurons.The Journal of Neurophysiology,1997,77(4):1994~2006
13 尼克尔斯(美)著,杨雄里等译.神经生物学:从神经元到脑.北京:科学出版社,2003(Nicholls,JG.Neurobiology:From neuron to brain(Yang X L,Trans.).Beijing:Science press,2003(in Chinese))
14 Rybak IA,Shevtsova N A,St-John W M,Paton JF,Pierre F O.Endogenous rhythm generation in the Pre-Bötzinger complex and ionic currents:modeling and in vitro studies.European Journal of Neuroscience,2003,18(2):239~257
15 Toporikova N,Butera R J.Two types of independent bursting mechanisms in inspiratory neurons:an integrative model.Journal of Computational Neuroscience,2011,30(3):515~528
16 Rybak I A,Paton J F,Schwaber J S.Modeling neural mechanisms for genesis of respiratory rhythm and pattern.II.Network models of the central respiratory pattern generator.The Journal of Neurophysiology,1997,77(4):2007~2026
猜你喜欢
安家(校外教育)(2022年6期)2022-01-03 11:47:06
军事文摘(2021年18期)2021-12-02 01:28:00
世界科学技术-中医药现代化(2021年7期)2021-11-04 08:10:42
北方音乐(2017年4期)2017-05-04 03:40:08
保健与生活(2017年5期)2017-02-27 00:32:17
祝您健康(2016年2期)2016-02-24 15:36:16
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国当代医药(2015年1期)2015-03-01 02:00:21
中国药理学通报(2014年2期)2014-05-09 08:22:33
中国输血杂志(2013年9期)2013-03-21 18:25:26
