
牡蛎幼虫光照实验提示复杂遗传背景下基因互作的多样性*
2015-03-08 08:48徐彩丽武长路于文超王晓通
海洋与湖沼 2015年3期
徐彩丽 许 飞 王 娇 武长路 于文超 王晓通①
(1. 鲁东大学农学院 烟台 264025; 2. 中国科学院海洋研究所 青岛 266071)
长牡蛎(Crassostrea gigas)在系统分类学上属软体动物门、双壳纲、牡蛎目、牡蛎科、巨蛎属, 味道鲜美, 营养价值高, 是人们喜食的海产佳肴, 素有“海中牛奶”之称, 是一种适合我国沿海养殖的优良品种, 经济效益显著, 现已成为我国重要的海水养殖品种(毛玉泽等, 2005)。很多研究表明, 葡萄牙牡蛎(Crassostrea angulata)也属于软体动物门、双壳纲、牡蛎目、牡蛎科、巨蛎属, 是长牡蛎的亚种, 在我国以长江为界, 主要分布在南方沿海, 在外形和生活习性方面与长牡蛎具有一定差异。
相对于品种之间, 亚种之间的遗传背景差距更大, 但已有证据表明两种牡蛎在自然条件下可以杂交, 且杂交后代具有正常的有丝分裂和减数分裂, 并且具有一定杂种优势(Soletchnik et al, 2002; Batista et al, 2007; 郑怀平等, 2012)。光照是水产养殖环境中的一项重要因素, 对于水产动物的生理状态具有一定影响, 曾有学者在室内试验条件下研究了光照对硬壳蛤稚贝的潜砂活动的影响, 结果显示光照对其潜砂活动有显著影响, 光照强度的增加能提高稚贝潜砂的比例(王国栋等, 2008); 还有学者观察到, 在有光照的情况下, 泥下蜗牛的摄食量会减少, 这与它一直生活在泥下环境的习性相一致(Magalhães et al,2012); 但光照对于牡蛎幼虫生长速度和成活率方面的研究还很少, 长牡蛎与葡萄牙牡蛎杂交幼虫对不同光照梯度的反应, 还未见报道。另外, 在牡蛎的育苗实践中, 大家仍未确定合适的光照强度, 还需要更多的实验证据。
本研究通过观察比较不同光照强度下长牡蛎幼虫及长牡蛎与葡萄牙牡蛎杂交幼虫壳高和成活率的差异, 揭示光照是否对牡蛎幼虫早期的生长成活率有影响、杂交牡蛎幼虫是否在这个时期是否表现出杂种优势, 为牡蛎幼虫的培养和杂交育种提供有参考数据。同时由于本研究中杂交牡蛎幼虫的父母本遗传背景差异较大, 而牡蛎本身的杂合度也很高, 基因互作的多样性是否会体现为杂交幼虫表型上的多样性,也需要进一步研究。
1 材料与方法
1.1 实验动物
长牡蛎采自山东乳山, 葡萄牙牡蛎采自福建莆田, 壳高均为8cm左右, 将牡蛎壳表面清洗干净, 放在育苗池中暂养。
1.2 实验仪器
主要包括充气泵、照度计、显微镜、水桶、光源、螺丝刀、洗瓶、烧杯、胚胎观察皿、碘酊等。
1.3 牡蛎雌雄的鉴别
用牡蛎刀将暂养 3d后的牡蛎的闭壳肌割断, 撬掉其右壳, 用消过毒的牙签在其性腺部位扎一下, 然后将牙签在事先准备好的滴在载玻片上的一滴清水中摇晃几下, 之后将载玻片放在显微镜下观察, 辨别牡蛎的性别。
1.4 受精
分别挑选出雄性长牡蛎、雌性长牡蛎和雌性葡萄牙牡蛎, 用牙签分别划破三种牡蛎性腺表皮, 动作要轻, 注意不要划破内脏团, 然后用洗瓶冲洗长牡蛎的精子长牡蛎卵子、葡萄牙牡蛎的卵子, 并分别收集于三个水桶内, 将长牡蛎的卵子和葡萄牙牡蛎的卵子中分别放入十分之一体积的精子, 使其结合受精,30min左右便可观察受精情况, 根据观察结果判断是否补加精子或洗卵, 培养22h左右至受精卵发育为D型幼虫。
1.5 光照梯度设置与幼虫养殖
将水箱中培养的幼虫摇匀后, 用1000mL的烧杯盛取500mL含有幼虫的海水, 实验设置3个生物学重复, 长牡蛎幼虫分别记为 GG1、GG2、GG3; 杂交幼虫分别记为GA1、GA2、GA3; 本实验共设置6个梯度, 分别为269、58、15、4、1.5和0 lx, 光照时间是每天早 8点至晚 8点, 共 12h; 装有长牡蛎幼虫的烧杯分别记为 GG1-1, GG1-2, ··, GG1-6; GG2-1, GG2-2, ··,GG2-6; GG3-1, GG3-2, ··, GG3-6; 装有杂交幼虫的烧 杯 分 别 记 为 GA1-1, GA1-2, ··, GA1-6; GA2-1,GA2-2, ··, GA2-6; GA3-1, GA3-2, ··, GA3-6; 将烧杯中的幼虫从 1日龄饲养到 10日龄, 每天用塑料吸管吸取1mL的小球藻喂养, 每天喂2次, 培养到10日龄, 即壳顶期幼虫期。
1.6 测量及数据统计
壳高的测量: 配合目镜测微尺和镜台测微尺进行壳高的测量, 选用带有刻度的塑料吸管吸取 1mL海水, 挤到胚胎观察皿中, 滴加一滴碘酊以免牡蛎幼虫移动而影响读数, 每个烧杯读30个样, 取均值。
成活率的计算: 用带刻度的塑料吸管分别在初始烧杯内吸取 1mL含有幼虫的海水, 挤到胚胎观察皿中, 滴加一滴碘酊以免牡蛎幼虫移动而影响读数,在显微镜下计算幼虫个数, 测三次取平均值, 然后再乘上烧杯中液体的体积, 即是烧杯中幼虫的初始总个数; 到第10天时, 按照同样的方法计算出第10天每个烧杯中幼虫的个数, 第 10天烧杯中幼虫的总个数与烧杯中幼虫的初始总个数的比值即为成活率;取样之前, 用塑料吸管多次吹打烧杯中的海水, 从而保证幼虫分布均匀。
1.7 数据分析与统计
使用 SAS软件的单因素最小二乘分析(GLM)对实验数据进行统计分析, 利用R软件的heatmap和pca程序对各家系进行聚类分析和主成分分析并制图。
2 结果
2.1 长牡蛎及杂交各家系幼虫在不同光照强度下的生长与成活情况
不同光照强度对长牡蛎和杂交牡蛎各家系的影响见表1和表2, 通过SAS软件的单因素最小二乘分析, 发现是长牡蛎各家系还是杂交牡蛎各家系, 各个光照梯度下第 10天的壳高均没有显著差异(P>0.05),而长牡蛎幼虫和杂交牡蛎幼虫均58 lx的光照强度下,表现出最高的成活率(卡方检验, P<0.05)。
2.2 不同光照强度对长牡蛎及杂交牡蛎各家系幼虫生长成活率影响的比较
将长牡蛎幼虫和杂交牡蛎幼虫在不同光照梯度下的壳高和成活率分别进行比较, 如图1和图2所示,长牡蛎幼虫和杂交牡蛎幼虫在各个光照梯度下的壳高均没有显著差异(P<0.05), 但杂交牡蛎幼虫的成活率在269 lx和58 lx两个光照梯度下均极显著的高于长牡蛎幼虫(P<0.01)。

表1 不同光照强度对长牡蛎生长和成活率的影响Tab.1 The shell height and survival rate of the Pacific oyster larvae under different illuminance levels

表2 不同光照强度对杂交牡蛎生长和成活率的影响Tab.2 The shell height and survival rate of the hybrid larvae from Pacific oyster and Portuguese oyster under different illuminance levels

图1 不同光照强度对长牡蛎及杂交牡蛎各家系幼虫生长速度影响的比较Fig.1 Comparison of the shell height between the Pacific oyster and the hybrid larvae from Pacific oyster and Portuguese oyster under different illuminance levels
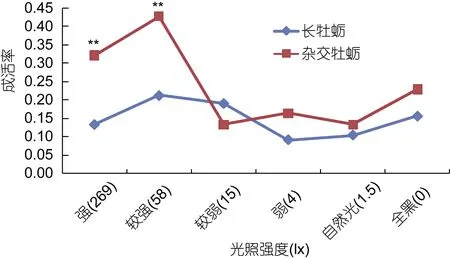
图2 不同光照强度对长牡蛎及杂交牡蛎各家系幼虫成活率影响的比较Fig.2 Comparison of the survival rate between the Pacific oyster and the hybrid larvae from Pacific oyster and Portuguese oyster under different illuminance levels
2.3 幼虫壳高与成活率相关性分析
为了验证是否因为饵料不足而造成壳高与成活率之间负相关, 我们将长牡蛎与杂交牡蛎的壳高与成活率合并在一起, 进行了相关性分析。如图3所示,R2=0.08远远小于 1, 表明壳高与成活率之间没有相关性, 壳高与成活率这两个指标各自独立, 不会对本文的分析结果造成不良影响。

图3 幼虫壳高与成活率相关性分析Fig.3 Correlation analysis of the shell height and the survival rate in all larvae
2.4 不同光照强度对同一母本的长牡蛎幼虫和杂交幼虫生长发育影响的比较
根据不同光照梯度下的壳高和成活率共12个指标, 对长牡蛎幼虫 3个家系(GG1、GG2、GG3)和杂交牡蛎幼虫 3个家系(GA1、GA2、GA3)进行聚类分析和主成分分析, 其中GG1与GA1家系来源于同一个雌牡蛎, GG2与 GA2家系来源于同一个雌牡蛎,GG3与GA3家系来源于同一个雌牡蛎。如聚类结果图4所示, 长牡蛎家系聚在一个大支上, 而杂交牡蛎家系GA1、GA2、GA3不能聚在一起, GA3位于长牡蛎家系的大支上, 而GA2、GA3分别位于不同的大支,且相隔距离较远。为了更形象地展示各个家系之间的差异, 作者又进行了主成分分析, 如图 5所示, 长牡蛎幼虫 3个家系(GG1、GG2、GG3)非常清晰的聚在一起, 相互之间间距不大, 而杂交牡蛎幼虫3个家系(GA1、GA2、GA3)很明显的分在三个方向, 相互之间间距都很大, 有意思的是, GG1与 GA1之间的距离、GG2与GA2之间的距离、GG3与GA3之间的距离都比较小, 这应该是因为母本是同一只牡蛎的原因。
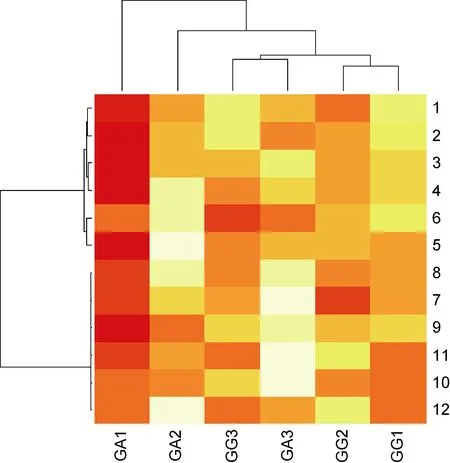
图4 不同光照强度对长牡蛎及杂交牡蛎各家系幼虫生长成活率影响的聚类分析Fig.4 Cluster analysis of the shell height and the survival rate of the Pacific oyster and the hybrid larvae from Pacific oyster and Portuguese oyster under different illuminance levels
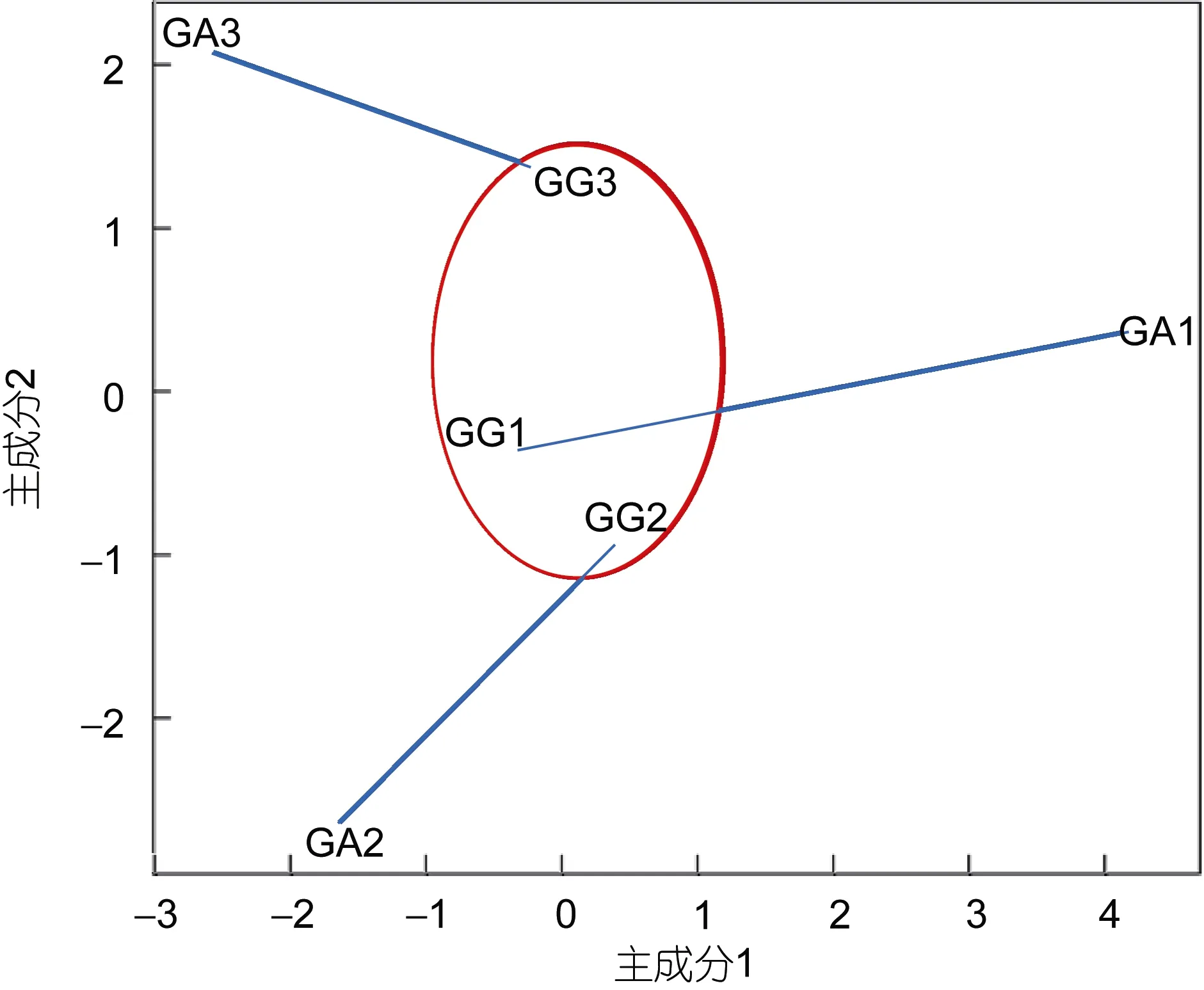
图5 不同光照强度对长牡蛎及杂交牡蛎各家系幼虫生长成活率影响的主成分分析Fig.5 Principal component analysis of the shell height and the survival rate of the Pacific oyster and the hybrid larvae from Pacific oyster and Portuguese oyster under different illuminance levels
3 讨论
光照是水产养殖环境中的一项重要因素, 光照强度对水生动物生长、发育及成活率的影响光照强度对水生动物生长的影响因种而异, 既能促进也能抑制(周显青等, 1999, 2000)。北极红点鲑(Satvetinus alpinus)幼体在50 lx时生长最快, 死亡率较低, 高于或低于此值的光照强度都会使生长速度减慢(Minagawa, 1994)。在本实验当中也有类似结果, 作者发现光照在0—269 lx范围内变化对牡蛎幼虫的生长没有显著影响, 但长牡蛎幼虫和杂交牡蛎幼虫均在58 lx的光照强度下, 表现出最高的成活率, 说明在牡蛎育苗的前10天如果保持这样的光照强度则对生产有利, 长牡蛎幼虫和杂交牡蛎幼虫的这种相似性,也支持葡萄牙牡蛎与长牡蛎是一个物种的观点(Wang et al, 2010; 林清等, 2014)。而杂交牡蛎幼虫在269 lx和58 lx两个光照梯度下的成活率均极显著的高于长牡蛎幼虫, 提示杂交牡蛎幼虫在这两个光照强度下表现出了杂种优势, 或许是因为长牡蛎与葡萄牙牡蛎的遗传背景具有较大差异但还没有产生生殖隔离,正好符合杂种优势产生的遗传条件(胡建广等, 1999;林建丽等, 2009)。
以不同光照梯度下壳高和成活率作为指标进行聚类分析和主成分分析后发现, 杂交牡蛎幼虫具有靠近各自同母本长牡蛎幼虫的趋势, 说明同一母本遗传背景在性状表现上启到了一定作用。但更有意思的是, 长牡蛎幼虫各家系间的表型比较接近, 而杂交牡蛎幼虫各家系之间差异非常大, 如果仅仅考虑基因序列的相似度, 3个杂交牡蛎幼虫的表型应该比较接近, 也应该聚在一起, 但事实是杂交幼虫表型的差异非常大, 作者推测是基因互作在杂交牡蛎的表型分化上发挥了重要作用, 牡蛎基因组的高杂合性(Zhang et al, 2012)使得长牡蛎和葡萄牙牡蛎各自储备了丰富的等位基因, 同时由于地理分布上的隔离,长牡蛎和葡萄牙储备的等位基因并不相同, 当两者杂交时, 两个亚种的不同基因座位的不同等位基因进入到了同一个个体中, 从而发生了亚种内很难发生的基因互作, 由于等位基因很丰富, 因此互作组合的类型也很多, 从而导致了表型的多样性。曾有学者发现杂交种在杂合状态下能够激发某种机制, 表达出与父母本均不同的新的基因产物, 调控相关的各种生理生化反应, 从而表现出杂种优势(王章奎等,2003; Baranwal et al, 2012; Fujimoto et al, 2012;Meyer et al, 2012), 本研究的发现也可以这样来解释,只是杂种优势的方向可能是多样化的。当然, 该推测还需更多的实验证据, 比如, 可以测定各组幼虫的转录组, 观察其对光照的反应模式是否与基因表达模式一致, 这是在今后研究中需要考虑的问题。
4 结论
根据长牡蛎及同母本杂交牡蛎幼虫对光照的不同反应, 作者认为, 58 lx的光照强度对前10天的牡蛎育苗生产有利; 葡萄牙牡蛎与长牡蛎的杂交是可行的, 在幼虫阶段就已经表现出了杂种优势; 具有高杂合度基因组并且遗传背景差异较大群体杂交一代表型的巨大差异可能由于基因互作组合多样化而引起。
王国栋, 张丽莉, 常亚青等, 2008. 温度, 光照和规格对硬壳蛤稚贝的潜砂活动的影响. 齐鲁渔业, 25(9): 3—5
王章奎, 倪中福, 孟凡荣等, 2003. 小麦杂交种及其亲本拔节期根系基因差异表达与杂种优势关系的初步研究. 中国农业科学, 36(5): 473—479
毛玉泽, 周 毅, 杨红生等, 2005. 长牡蛎(Crassostrea gigas)代谢率的季节变化及其与夏季死亡关系的探讨. 海洋与湖沼, 36(5): 445—451
林 清, 王亚骏, 王迪文等, 2014. 太平洋牡蛎和葡萄牙牡蛎养殖群体数量性状比较分析. 海洋通报, 33(1): 106—111
林建丽, 朱正歌, 高建伟, 2009. 植物杂种优势研究进展. 华北农学报, 24(S2): 46—56
周显青, 牛翠娟, 李庆芬, 1999. 光照对水生动物行为的影响.动物学杂志, 19(2): 45—48
周显青, 牛翠娟, 李庆芬, 2000. 光照对水生动物摄食、生长和存活的影响. 水生生物学报, 24(2): 178—181
郑怀平, 王迪文, 林 清等, 2012. 太平洋牡蛎与葡萄牙牡蛎两近缘种间杂交及其早期阶段生长与存活的杂种优势.水产学报, 36(2): 210—215
胡建广, 杨金水, 1999. 作物杂种优势的遗传学基础. 遗传,21(2): 47—50
Baranwal V K, Mikkilineni V, Zehr U B et al, 2012. Heterosis:emerging ideas about hybrid vigour. Journal of Experimental Botany, 63(18): 6309—6314
Batista F M, Leito A, Fonseca V G et al, 2007. Individual relationship between aneuploidy of gill cells and growth rate in the cupped oysters Crassostrea angulata, C. gigas and their reciprocal hybrids. Journal of Experiment, 352(1):226—233
Fujimoto R, Taylor J M, Shirasawa S et al, 2012. Heterosis of Arabidopsis hybrids between C24 and Col is associated with increased photosynthesis capacity. Proceedings of the National Academy of Sciences USA, 109(18): 7109—7114
Magalhães T R, Neves R A, Valentin J L et al, 2012. Do the changes in temperature and light affect the functional response of the benthic mud snail Heleobia australis(Mollusca: Gastropoda)? Anais Da Academia Brasileira De Ciencias, 86(3): 1197—1206
Meyer R C, Witucka-Wall H, Becher M et al, 2012. Heterosis manifestation during early Arabidopsis seedling development is characterized by intermediate gene expression and enhanced metabolic activity in the hybrids.The Plant Journal, 71(4): 669—683
Minagawa M, 1994. Effects of photoperiod on survival, feeding and development of larvae of the red frog crab, Raninaranina.Aqaculture, 120(1—2): 105—l14
Soletchnik P, Huvet A, Moine O L et al, 2002. A comparative field
study of growth, survival and reproduction of Crassostrea gigas, C.angulata and their hybrids. Aquatic Living Resources, 15(4):243—250
Wang H Y, Qian L M, Liu X et al, 2010. Classification of a common cupped oyster from southern China. Journal of Shellfish Research, 29(4): 857—866
Zhang G, Fang X, Guo X et al, 2012. The oyster genome reveals stress adaptation and complexity of shell formation. Nature,490(7418): 4954
猜你喜欢
发明与创新(2022年36期)2022-12-02
蔬菜(2021年7期)2021-11-27
环球时报(2017-12-25)2017-12-25
大众考古(2015年6期)2015-06-26
红领巾·探索(2014年7期)2014-10-10
种子科技(2014年8期)2014-05-18
小说月刊(2014年8期)2014-04-19
中学生数理化·八年级物理人教版(2014年2期)2014-04-02
种子科技(2014年9期)2014-01-22
西部(2013年3期)2013-11-16
