
2013年云南省禽流感病毒H9N2亚型监测及其血凝素基因序列分析
2015-03-08 09:35胡媛媛张文东宋建领刘庆亮郑锦玲保志鹏张应国范泉水赵焕云张富强
中国预防兽医学报 2015年1期
胡媛媛,张文东,宋建领,刘庆亮,曾 伟,王 锐,郑锦玲,彭 洁,保志鹏,张应国,范泉水,赵焕云*,张富强*
(1.云南农业职业技术学院,云南 昆明 650031;2.云南省动物疫病预防控制中心,云南 昆明 650051;3.云南省热带亚热带动物病毒病重点实验室,云南 昆明 650224;4.云南农业大学,云南 昆明 650223;5.成都军区疾病预防控制中心,四川 成都 100021)
H9N2 亚型为低致病性禽流感病毒(Avian influenza virus,AIV),可引起家禽呼吸困难、腹泻、产蛋下降等症状,当继发细菌感染时可引起较高的死亡率。我国自1994 年首次从鸡群中分离到H9 亚型HA 以来[1],绝大部分省市均存在H9 亚型AIV 的流行,给我国养禽业造成严重经济损失。1998 年后又先后从猪和人体内分离到H9N2 AIV[2]。AIV 属于正粘病毒科流感病毒属A 型流感病毒成员,病毒基因组由单股、负链、8 节段RNA 组成,含有10 个基因,其中第4 和第6 节段RNA 分别编码血凝素(HA)和神经氨酸酶(NA)。目前已知HA 共有16 种亚型(H1~H16)、NA 共有9 种亚型(N1~N9)[3]。HA是流感病毒主要表面抗原和保护性抗原,HA 与细胞表面受体的相互作用,介导病毒吸附和侵入细胞的过程,而其活性依赖N 末端信号肽的切除及HA1和HA2 的产生。HA 的裂解位点、糖基化位点、受体结合位点等关键性氨基酸的突变是病毒抗原性和致病性变异的分子基础之一[4-5]。
为分析云南地区H9N2 亚型AIV 相关致病性基因变异情况,本研究对2013 年检测到的部分H9N2亚型AIV 阳性样品中的HA 基因进行了测序比对分析,表明云南地区流行的AIV 存在一定的变异,本研究结果为禽流感防控及制苗种毒更新提供了实验依据。
1 材料和方法
1.1 病毒株、质粒及临床样品 灭活H9N2 亚型AIV(作为检测的阳性对照)购自哈尔滨维科生物技术开发公司;pMD18-T 购自TaKaRa 公司;3 622 份临床样品为2013 年采自云南省活禽交易市场、规模场、散养户等的家禽咽喉棉拭子样品。
1.2 主要试剂 MiniBEST Viral DNA/RNA Extraction Kit、One Step RNA PCR Kit(AMV)、Premix Taq 酶、DL2000 DNA Marker 购自TaKaRa 公司;DNA 凝胶胶回收试剂盒、小量质粒纯化试剂盒购自爱思进生物技术(杭州)有限公司。
1.3 H9N2亚型RT-PCR 及多重RT-PCR 检测按试剂盒提供的操作手册,从接种采集的临床样品的鸡胚尿囊液中提取总RNA。采用H9N2 亚型特异性RT-PCR 及多重RT-PCR 进行检测[6]。
1.4 HA基因的扩增 采用文献[7]的方法,以多重RT-PCR 鉴定为阳性的核酸样品为模板,利用特异性引物(5'-CACCATGGAAACAATATATCACTAM T-3'/5'-CTATATACAAATGTTGCRYCT-3')扩增病毒HA 基因。预期扩增片段为1 687 bp,编码560 aa。
1.5 序列同源性比对及系统进化分析 将PCR 产物克隆于pMD18-T 载体中,由深圳华大基因科技服务有限公司进行测序。采用MEGA6 软件进行AIV分离株HA 基因序列的比对和系统进化分析。所引用的代表病毒株序列来自GenBank(表1)。
2 结果
2.1 H 9N2亚型RT-PCR 及多重RT-PCR 检测采用H9N2 亚型RT-PCR 对检测样品进行扩增,扩增产物分别约为310 bp 和170 bp,与预期结果相符。2013 年云南省16 个地区的3 622 份家禽样品中,共分离检测出H9N2 亚型AIV 阳性97 份,每个地区选取1~3 份时间、地点、宿主有差异的分离株,共21 份,进行HA 基因的克隆、测序。
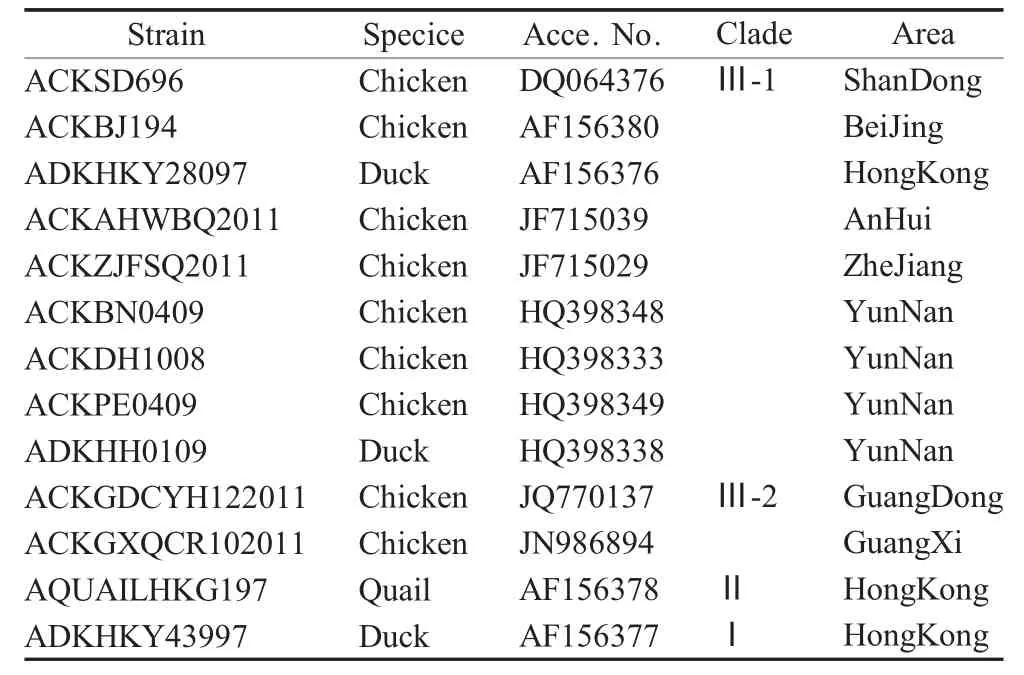
表1 序列比对及系统进化分析中引用的H9N2 亚型参考病毒株Table 1 Reference strains of H9N2 subtype virus used in sequence alignment and phylogenetic analysis
2.2 序列同源性及系统进化分析 将21 个分离株中H9N2 亚型AIV HA 基因序列与参考病毒株序列进行同源性比较及系统进化分析。结果显示21 个H9N2 亚型病毒株HA 基因核苷酸同源性为86.0 %~99.4 %,与云南省现用H9N2 亚型疫苗株ACKSD696 同源性为89.2 %~91.0 %;与欧亚分支中的类A/Chicken/BeiJing/I/1994(分支Ⅲ)分支代表株ACKBJ194 和ADKHKY28097 亲缘关系比较近,为89.5%~92.2%,与欧亚分支中的其它2 个分支(类A/Quail/Hong Kong/G1/97(分支Ⅱ)、类A/Duck/Hong Kong/Y439/97(分支Ⅰ)代表病毒株亲缘关系较远,同源性分别为85.5%~87.5%、80.6%~82.9%;2013年部分云南分离株与2008 年~2009 年云南分离株的同源性关系相对较远,同源性为86.9 %~94.8 %,大部分云南病毒株与2011 年华南代表株ACKGD 122011 及ACKGX102011 同源关系比较近,同源性为86.3 %~98.2 %,其余少部分分离株与2011 华东及华中代表株ACKAH2011 和ACKZJ2011 同源关系比较近,同源性为96.3 %~98.0 %。
系统进化分析表明2013 年云南H9N2 亚型AIV属于欧亚谱系中的类A/Chiken/BeiJjing/I/1994 分支(分支Ⅲ),并且又进一步划分为2 个不同的小分支(Ⅲ-1 和Ⅲ-2),其中Ⅲ-1 分支的6 个分离株与华东和华中株同属于一个分支,Ⅲ-2 分支的15 个分离株与华南株同属一个分支(图1)。
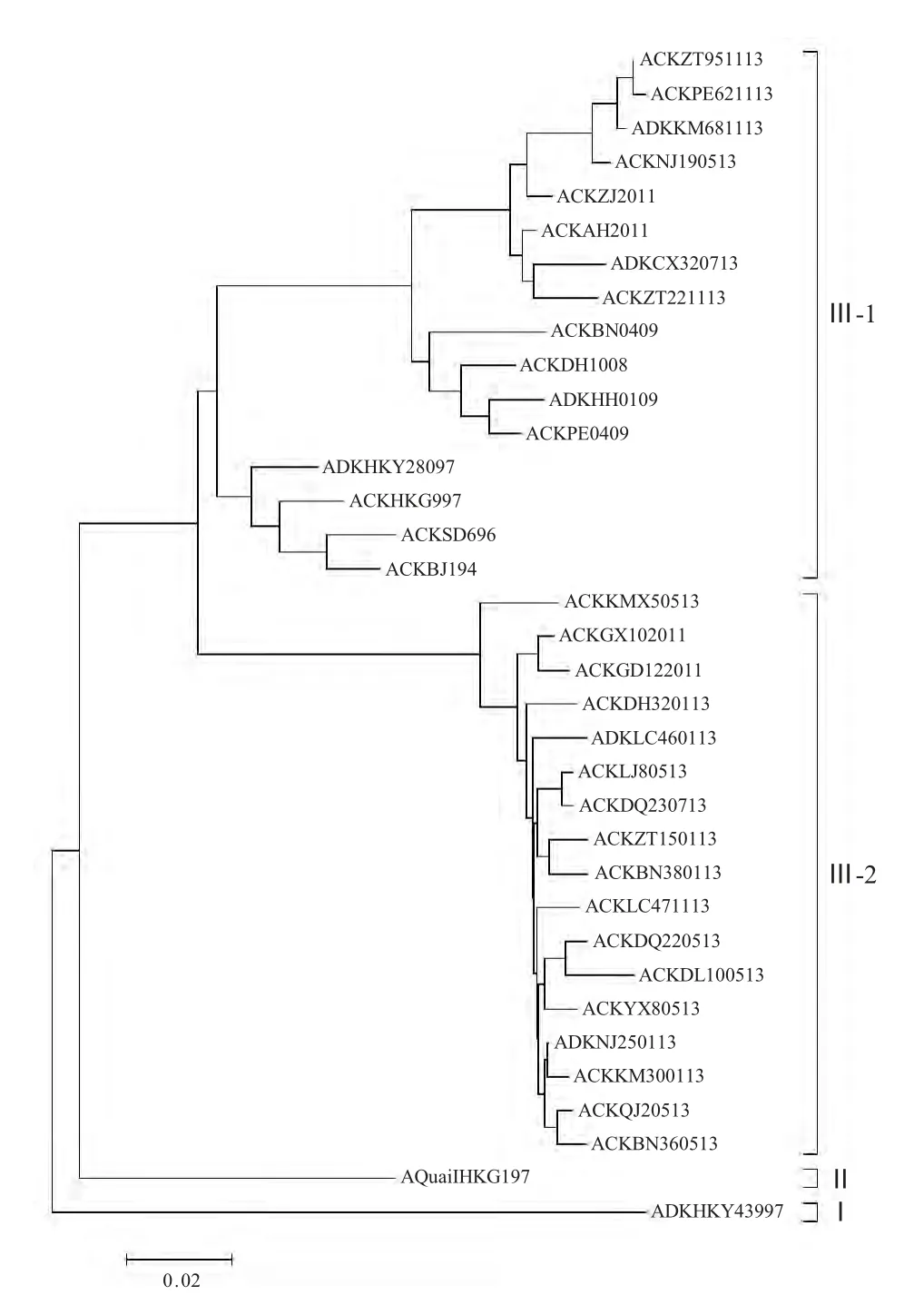
图1 2013 年云南省H9N2 亚型AIV HA 基因系统发育分析Fig.1 Phylogenetic analysis based on HA genes of H9N2 subtype AIVs from Yunnan province in 2013
2.3 氨基酸序列比对 推导氨基酸序列分析结果显示,测序的21 个HA 氨基酸序列与疫苗株ACKSD696 比较,有多个位点存在差异,其中HA存在Q90M(W)的一致性差异。此外,分离株Ⅲ-1 和Ⅲ-2 两个分支间有21 个氨基酸存在差异,Ⅲ-1 分支 在15、63、127、181、216、253、283、285、287、334 位氨基酸位点存在特征性变异,Ⅲ-2 分支在15、88、107、138、166、168、171、243、269、370、441 氨基酸位点存在特征性变异,其中Ⅲ-1 分支中ACKNJ190513、ADKKM681113、ACKPE621113及ACKZT951113 分离株在89、130、164、201、204、451 出现特有的变异(表2)。
所有分离株的HA 裂解位点均为RSSR↓GLF,具有低致病性病毒分子特征[8]。与公认的aa29~aa31、aa141~aa143、aa218~aa220、aa298~aa300、aa305~aa307、aa492~aa494、aa551~aa553 7 个潜在的糖基化位点相比较,只有ADKCX320713 具有全部糖基化位点,其余分离株均有变化,其中除ADKCX320713 和ACKZT221113 在aa218 位点存在糖基化位点外,其余均为T218I 点突变导致其糖基化位点消失。ACKKMX50513、ACKBN380113 和ACKZT221113 分别因N141S、T298A、N551K 点突变失去一个糖基化位点;Ⅲ-1 分支病毒株在313 位出现了一个潜在的糖基化位点。HA 受体结合位点大部分均未发生变化,只有在第146、191、198、234、235 位氨基酸存在变异,其中第234 位氨基酸全部变为L,呈现了人流感受体结合特性;Ⅲ-1 分支分离株在A198V(T)和Q235M(W)存在变异(表3)。
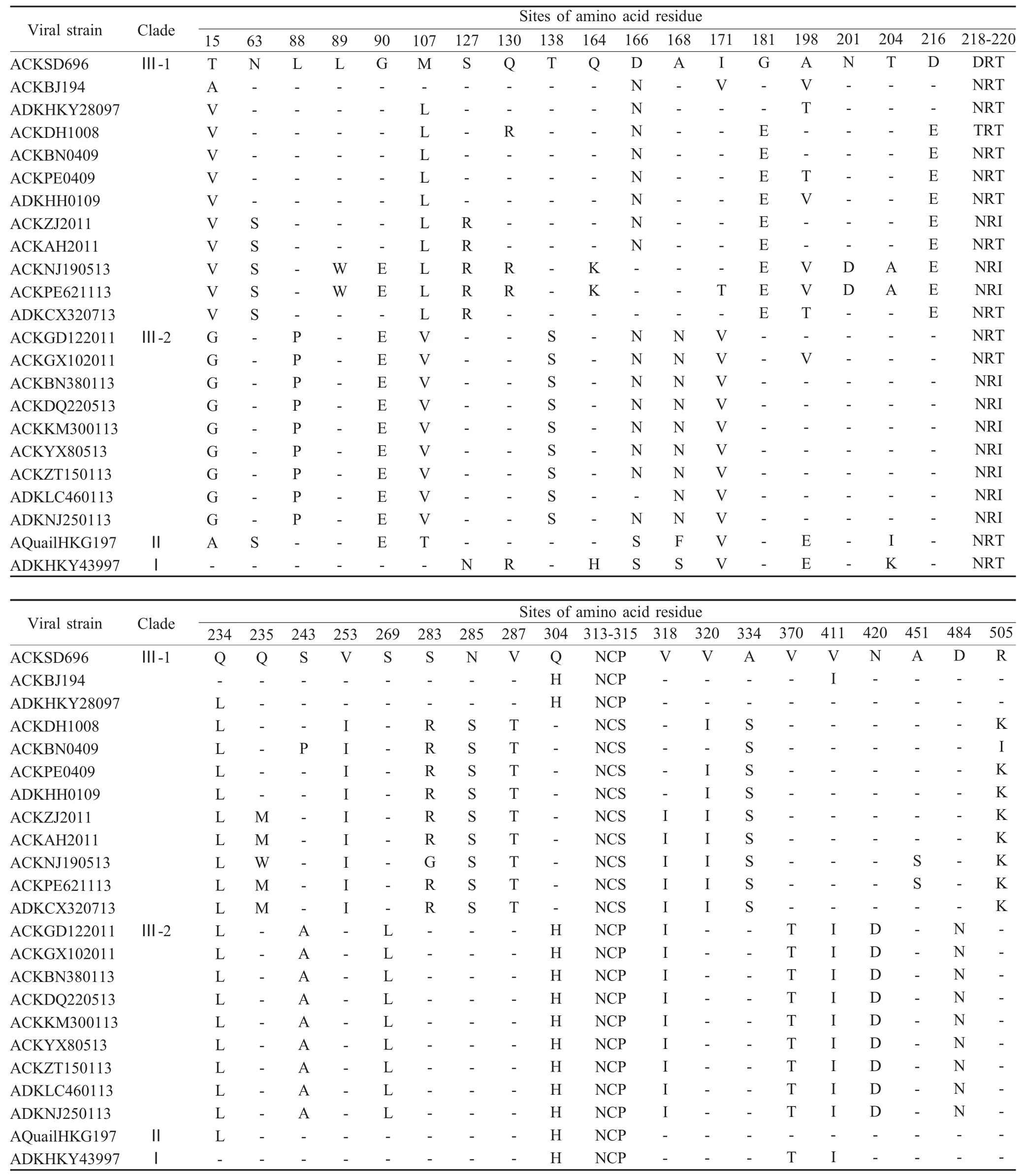
表2 2013 年云南省AIV H9N2 亚型HA 氨基酸突变位点比对分析Table 2 Alignment of other mutant amino acid sites on HA of H9N2 viruses

表3 2013 年云南省AIV H9N2 亚型抗原表位关键性氨基酸位点比对Table 3 Alignment of antigenic epitope sites on HA of H9N2 viruses
3 讨论
系统进化分析表明2013 年云南H9N2 亚型AIV分离株既具有华中华东株遗传特征,又具有华南株遗传特征,并且每个分支病毒株分离的时间和地域分布,并无明显差异,表明云南省H9N2 亚型AIV来源比较复杂,这是否与云南鸡雏来源有关还有待于进一步的调查。云南省现用疫苗株为96 年山东株(ACKSD696),同源性分析表明2013 年云南分离株与其亲缘关系较远,需要密切关注当前疫苗的免疫保护情况。此外,Ⅲ-1 分支为云南省2008 年~2009年H9N2 亚型病毒株的延续;Ⅲ-2 分支为云南省新出现的进化分支。针对2013 年AIV H9N2 亚型HA氨基酸位点比对发现,Ⅲ-1 分支中2008 年以后出现的病毒在公认的7 个糖基化位点均发生氨基酸替代,并导致aa313~aa315 出现了一个新的潜在糖基化位点;2011 年以后出现的病毒aa63、aa127、aa235,部分2013 年出现的病毒aa89、aa130、aa164、aa201、aa204、aa451 均发生氨基酸替代。Ⅲ-2 分支中病毒HA aa88、aa107、aa138、aa168、aa243、aa269、aa420、aa484 位点均发生氨基酸替代。Ⅰ分支及Ⅲ-2 分支病毒HA aa370、aa401,Ⅲ-2分支及部分2013 年Ⅲ-1 分支病毒aa90,所有2008年及以后在云南出现的病毒及参考株aa15,2011 年及以后出现的病毒在aa318 位点均存在氨基酸替代。这种替代具有一定的进化(亚)分支特异性,是否会影响病毒的生物学特性还有待进一步的研究。
2013 年云南H9N2 分离株aa234 位点全部变为L,呈现了人流感受体结合特性,可能会进一步对人类公共卫生造成威胁。有报道称HA 的受体结合位点中aa191 位点的氨基酸最保守[9],2013 年云南分离株与已报道的HA 蛋白抗原表位关键性氨基酸位点相比较,Ⅲ-1 分支中部分2013 年分离株、Ⅰ和Ⅱ分支在aa153 位点发生氨基酸替代,Ⅲ-1 分支中部分2013 年病毒株在aa201 位点发生氨基酸替代。Ⅲ-2 分支在aa143 位氨基酸与疫苗株一致,但与部分Ⅲ-1 分支、Ⅰ和Ⅱ分支不同。表明虽同属一个进化分支,但由于抗原表位关键性氨基酸不同,可能其抗原性也有所不同。
[1]陈伯伦,张泽纪.禽流感研究I:鸡A 型禽流感病毒的离与血清学初步鉴定[J].中国兽医杂志,1994,22(10):3-5.
[2]李曦,于康震,田国斌,等.H9N2 亚型禽流感病毒血凝素基因序列的初步分析[J].黑龙江畜牧兽医,2003,2:29-30.
[3]List A and B diseases of mammals,birds and bees:manual of standards for diagnostic tests and vaccines[M].Office International des Epizootites,2004,5th Edition.
[4]Nikolai V K,Irina A R,Natalia A I,et al.Structure of antigen-ic sites on the haemagglutinin molecule of H5 avian influenza v-irus and phenotypic variation of escape mutants[J].J Gen Virol,2002,83:2497-2505.
[5]Wai Lan-wu,Lin Tian-wei,Xia Ning-shao,et al.Antigenic profile of avian H5N1 viruses in Asia from 2002 to 2007[J].J Virol,2008,82(4):1798-1807.
[6]张应国,宋建领.禽流感病毒RT-PCR 及多重RT-PCR 检测技术的建立[J].中国兽医科技,2005,35(08):600-604.
[7]宋建领,叶丛华,田建国,等.云南2008~2009 年H9N2亚型禽流感病毒血凝素和神经氨酸酶基因序列分析[J].畜牧与兽医,2011,43(3):48-53.
[8]Liu Hong-qi,Liu Xiu-fan,Cheng Jian,et al.Phylogeneti analysis of the hemagglutinin genes of twenty-six avian influenza viruses of subtype H9N2 isolated from chickens in China during 1996-2001[J].Avian Dis,2003,47:116-127.
[9]贾红玲,苏艳,吴润.禽流感病毒血凝素保守氨基酸对其受体结合位点的影响[J].中国预防兽医学报,2008,30(7),500-504.
猜你喜欢
黑龙江大学自然科学学报(2022年1期)2022-03-29
计算机系统应用(2021年10期)2022-01-06
复旦学报(医学版)(2021年5期)2021-10-13
学生天地(2019年28期)2019-08-25
癌症进展(2018年11期)2018-12-30
环球时报(2018-03-19)2018-03-19
北方牧业(2016年15期)2016-12-17
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
小星星·阅读100分(高年级)(2014年6期)2014-06-27
