
产毒与不产毒铜绿微囊藻对模拟酸雨及紫外辐射的生理响应
2015-02-11 02:06杨雨玲董丽丽李亚鹤高坤山
生态学报 2015年23期
李 伟,杨雨玲,黄 松,董丽丽,潘 健,李亚鹤,周 月,高坤山
1 黄山学院生命与环境科学学院,黄山 245041 2 厦门大学近海海洋环境科学国家重点实验室,厦门 361005 3 宁波大学海洋学院,宁波 315211
产毒与不产毒铜绿微囊藻对模拟酸雨及紫外辐射的生理响应
李 伟1,2,杨雨玲1,黄 松1,董丽丽1,潘 健1,李亚鹤2,3,周 月1,高坤山2,*
1 黄山学院生命与环境科学学院,黄山 245041 2 厦门大学近海海洋环境科学国家重点实验室,厦门 361005 3 宁波大学海洋学院,宁波 315211
为了比较研究酸雨与紫外辐射对淡水水体常见藻华蓝藻的生理学影响,选取铜绿微囊藻(Microcystisaeruginosa)产毒(FACHB-905)与不产毒(FACHB-469)株系作为实验材料,通过人工模拟酸雨,研究了不同pH处理后2藻株的光合生理变化以及对紫外辐射的敏感性的异同。实验设置3个pH梯度,pH7.10为对照组(正常培养基培养的藻体),两模拟酸雨处理组(pH5.65和pH4.50);两种辐射处理,可见光处理(PAR)以及全波长辐射处理(PAB)。研究结果表明,905藻株细胞粒径在各pH处理下都要显著高于469藻株,模拟酸雨处理显著降低了两藻株细胞的平均粒径及体积,但叶绿素含量显著提高;酸雨处理同时也引起细胞死亡率的增加,表现为藻体有效光化学效率显著降低,生长速率显著受到抑制,低pH下呈负增长,且这种抑制程度在469下更为显著。高的可见光以及紫外辐射处理,使两株系有效光化学效率随pH的降低而呈降低趋势,其中469藻株降低至更低的水平,且高光辐射以及紫外诱导的抑制率要显著高于905藻体,这可能与469藻株较低的光保护色素有关(较低的类胡萝卜素以及紫外吸收物质)。在未来全球变化背景下,不同种类的浮游植物对环境变化的响应及适应能力不同,可改变水体的群落结构和种群丰度,铜绿微囊藻905较469较强的耐受酸雨以及紫外辐射的能力,可能会使该株系在竞争力上占据优势。
铜绿微囊藻;酸雨;紫外辐射;生长;光合特性
酸雨是继温室效应、臭氧空洞等环境问题之后的第三大环境问题[1],酸雨引起的潜在环境影响以及经济损失已经成为世界各国关注的热点问题[2]。已有研究表明,酸雨导致的水体H+及Al3+浓度增加可引起对酸性敏感的鱼类的离子、渗透压调节以及气体交换功能的紊乱[3-4],并可与酸性水体中阳离子(钙、镁、钠等)的丢失协同影响水生动物的生理、代谢活动[5-6]。在浮游植物中,酸雨可影响水体浮游植物叶绿素含量[7],改变浮游植物的光合生理以及群落结构,低pH下水生浮游植物多样性显著受到抑制[8-9]。
阳光紫外辐射(UVR)可对大多数水生浮游植物产生负面影响。UVR可降低浮游植物的光合效率,抑制浮游植物的生长[10],并可与其他环境因子,如营养盐[11]、光强[12]等耦合降低浮游植物的初级生产过程。在当前气候变化背景下,UVR强度的增加以及区域性的分布不均衡[13-14],将会协同其他环境因子对水生浮游植物的光合生理产生长期、持续的影响[15],且水体酸化导致水体溶解有机质含量降低,可进一步增强UV辐射在水体中的强度和透射深度[16]。
在应对环境胁迫时,不同种类浮游植物的响应、适应能力具有较大差异[17],这种差异性可改变水体的浮游植物群落结构以及种群丰度。蓝藻作为最古老的一类光合自养生物,对环境变化的适应能力较强[18]。虽然有关蓝藻(螺旋藻、微囊藻、念珠藻等)对UV辐射的响应得到了一定的研究[19-21],但很少涉及酸雨与紫外辐射耦合对典型蓝藻生理、生化的影响,尤其是选取具有代表性的水华蓝藻铜绿微囊藻产毒与不产毒株进行比较性研究,尚未见报导。通过对以往原位调查的研究可知,自然界中常见藻华种类多以危害较大的产毒株为主[22],但具体形成机制如何尚需进一步探讨。不同类型的微囊藻光合生理在响应主要环境变化(酸雨、UV辐射等)的能力上是否具有差异,以及差异程度如何,可能是决定未来哪种藻在竞争地位上更占优势的主要因素。因此,本研究选取我国重要的淡水水华蓝藻铜绿微囊藻的两个不同品系为研究对象,希望从生理学的角度探讨酸雨对两株藻光合生理的影响以及二者响应的异同,并且研究经酸雨处理后两株藻体对紫外辐射的敏感性和耐受能力,以期了解不同类型浮游植物响应环境变化的生理学差异,为进一步探究微囊藻水华的优势类群提供生理学数据,同时也能够为预测未来微囊藻水华发生的优势种类提供理论依据。
1 材料方法
1.1 实验材料及培养条件
铜绿微囊藻FACHB-905(产毒素)及FACHB-469(不产毒素)由马增岭副教授(温州大学) 惠赠。藻体置于生物培养箱内使用BG11(+N)培养基培养,温度为20℃,培养光强为120μmol m-2s-1,光暗比(L∶D)为12∶12,在此条件下适应培养20代(指数增长期)后进行实验。
1.2 实验方法1.2.1 模拟酸雨及pH值的测定
模拟酸雨母液使用98%的H2SO4和65%HNO3按照摩尔比8∶1进行配制。通过向已经加入BG11(+N)培养基的溶液中加入不同体积的上述酸雨母液,获得3个pH梯度,即pH 7.10(对照组,正常培养基不加模拟酸雨母液)以及模拟酸雨处理组pH 5.65和pH 4.50。模拟不同pH的酸雨过程中,培养基的pH测定使用Mettler Toledo DL15 Titrator(Sweden)进行,在测定前,使用NBS(National Bureau of Standards)缓冲液(pH 4.01,7.01和pH 10.01)进行校正。
1.2.2 藻细胞模拟酸雨处理
将正常培养状态下的藻细胞(指数增长期)转移至提前调节好pH的模拟酸雨培养基中,初始细胞浓度为每毫升5×104个细胞,置于体积为1L的聚碳酸酯瓶中密闭培养24小时,除pH7.10下细胞浓度增加,低pH处理下细胞浓度增长较少或呈负增长,在此条件下,pH值在培养期间变动较小,整体保持稳定水平(表1)。培养光照、温度及光暗比同上。培养期间定期摇晃培养瓶以保证细胞充分悬浮。每个处理下3个重复培养。

表1 铜绿微囊藻FACHB-469和FACHB-905经不同pH处理前后培养体系的pH值Table 1 The pH values in Microcystis aeruginosa FACHB-469 and FACHB-905 at the initial and after 24 hours culture in different pH treatments
1.2.3 比生长速率的测定
比生长速率(μ)的测定根据公式求得:
μ=(lnNt-lnN0)/t
式中,Nt为各pH下处理24h(t)后的细胞浓度,N0为初始细胞浓度。各处理下藻体细胞浓度使用Z2 Coulter Counter(Beckman Instruments, Florida, US)进行计数,通过该仪器可获得藻体细胞粒径以及细胞体积分布图。
1.2.4 色素及紫外吸收物质(UVACs)的测定
将经不同pH处理的藻体过滤至Whatman GF/F滤膜(孔径0.22 μm),置于15 mL离心管中,加入5mL甲醇后放入4℃冰箱内过夜提取(12h),然后将提取液在离心机内离心10min(5000r/min),最后用分光光度计测定上清液全波段(250—700nm)的吸光值。
叶绿素a含量的测定参照Porra[23]的方法进行计算:
叶绿素a (μg/mL)=16.29×(A665-A750)-8.54×(A652-A750)
类胡萝卜素含量的测定参照Parsons & Stricklan[24]的方法进行计算:
类胡萝卜素 (μg/mL)=7.6×[(A480-A750)-1.49×(A510-A750)]
紫外吸收物质(UVACs)的含量测定参照徐军田的计算方法进行,即通过计算UVACs吸收峰的峰高值(H)与藻体叶绿素a含量的比来表示UVACs的含量[25]。
1.2.5 紫外辐射处理
将不同pH下培养24h后的藻体(905、469)分装至石英管(可透UVR)。每个pH水平下的藻体分别给予两种辐射处理即:(1)可见光 (PAR)处理(400—700nm),在石英管外包裹一层Ultraphan395 滤膜(UV Opak, Digefra,德国慕尼黑),滤除紫外线A(UV-A,320—400nm)和紫外线B(UV-B,280—315 nm);(2) 可见光+紫外系A+紫外线B(PAB)处理(295—700nm),在石英管外包裹一层Ultraphan 295 滤膜(UVOpak, Digefra, 德国慕尼黑生产),该膜可以使藻体接受全波段的辐射。每种辐射处理下3个重复。整个辐射处理在人工光源太阳模拟器 (Sol 1200W; A. G. Hönle, Martinsried, Germany)下进行,通过调整石英管与太阳模拟器间的距离,调节辐射强度。本实验的光强设置分别为,PAR的强度为87.5 W/m2,UV-A的强度为33.5 W/m2,UV-B的强度为1.91 W/m2(参考我国南方地区夏季UVB强度设置)。
1.2.6 叶绿素荧光参数测定
将培养光强下的藻体直接取样,在氙灯脉冲调制荧光仪(XE-PAM, Walz,德国)下测定培养状态下的有效光化学效率Yield值。将各pH下处理24h后的藻体置于太阳模拟器下进行可见光(PAR)及可见光加紫外线A加紫外线B(PAB)辐射处理1h,处理过程中每隔15min对其Yield进行测定,共测定4轮。其中饱和脉冲光强设定为5000μmol m-2s-1,饱和脉冲持续时间设定为0.8 s。
Yield的计算方法如下:
Yield = Fm′- F0/ Fm′
式中,Yield为光系统Ⅱ的有效光化学效率,F0为本底叶绿素荧光,Fm′为在预设的光化光背景下的最大叶绿素荧光。
1.2.7 数据统计分析
数据处理使用Origin 7.0和SPSS 16.0软件,显著性分析使用one-way ANOVA(Turkey),two/three-way ANOVA用于分析因子间的相互作用,显著性水平P设置为0.05。
相对于初始的Yield,阳光模拟器下PAR及PAB导致的光化学效率Yield的抑制率计算方式如下:
lnhPAR= (PInitial-PPAR)/PInitial× 100%
lnhPAB= (PInitial-PPAB)/PInitial× 100%
式中,lnhPAR及lnhPAB为照射PAR及PAB 1h后相对于PInitial(初始Yield值)的抑制率,PPAR及PPAB为PAR及PAB下的Yield。
2 结果
2.1 色素
铜绿微囊藻905叶绿素a和类胡萝卜素含量随模拟酸雨处理呈增加趋势,相对于pH7.10处理,pH5.65处理下藻体叶绿素a及类胡萝卜素含量显著升高了11.33%(P<0.05)和4.22%(P<0.05),pH 4.50处理下显著升高了61.26%(P<0.05)和20.75%(P<0.05),紫外吸收物质UVACs在各pH处理下无显著差异(P>0.05)(图1)。
铜绿微囊藻469叶绿素a含量在低pH(pH 4.50)下显著增加了20.95%(P<0.05),而类胡萝卜素含量显著降低了67.5%(P<0.05),3个pH处理下紫外吸收物质含量差异显著(P<0.05),其中pH5.65处理下最高(0.44±0.04),pH4.50处理下最低(0.10±0.02)(图1)。
对比两藻种间色素水平,除pH7.10下叶绿素a含量在两藻株间无显著性差异外(P>0.05),其余pH处理下,叶绿素a、类胡萝卜素及紫外吸收物质含量在铜绿微囊藻905中含量都显著高于469株(P<0.05)。pH5.65下,905株系叶绿素a、类胡萝卜素及紫外吸收物质含量显著高于469株系8.36%(P<0.05),34.00%(P<0.05)和153.29%(P<0.05);pH 4.50下,则分别提高了0.201(P<0.05),3.30(P<0.05)和11.23(P<0.05)倍(图1)。

图1 铜绿微囊藻469与905在pH 7.10以及模拟酸雨(pH5.65及pH4.50)处理下24h后叶绿素a、类胡萝卜素以及紫外吸收物质含量(n=3)Fig.1 Chl a, carotenoid and UVACs of Mycrocystis aeruginosa strains of 469 and 905 after cultured under pH 7.10and simulated acid rain (pH5.65 and pH4.50) condition for 24h (n=3)
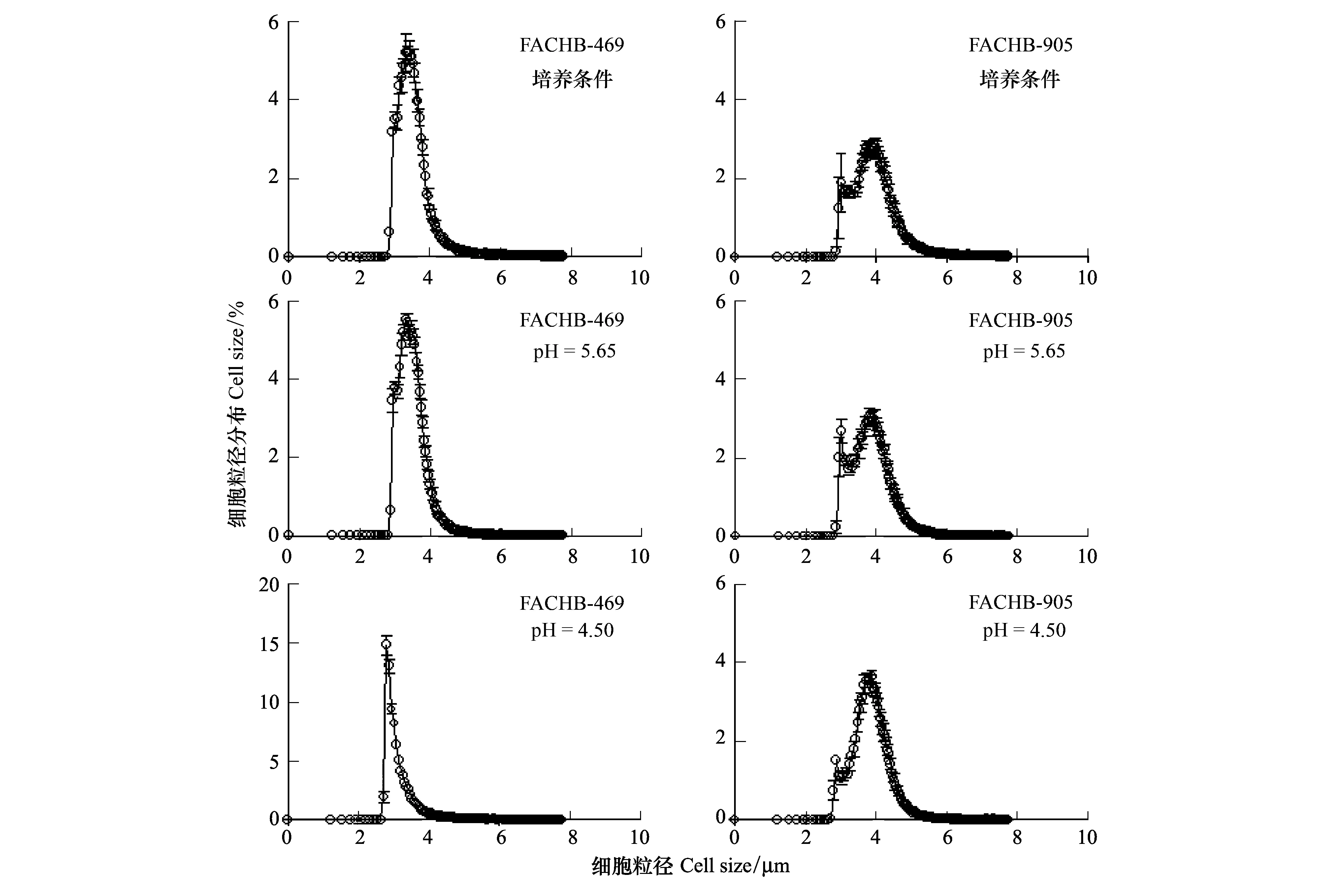
图2 铜绿微囊藻469 (FACHB-469)与905 (FACHB-905)在培养条件下以及模拟酸雨pH为 5.65及4.50下24h后细胞粒径分布图(n=3)Fig.2 The cell size of Mycrocystis aeruginosa strains of 469 and 905 under culture condition and after treated with simulated acid rain at pH of 5.65 and 4.50for 24h (n=3)
2.2 细胞粒径和体积
正常培养条件下(pH7.10),铜绿微囊藻469和905的细胞粒径和体积差异显著(图2 A, B),平均粒径分别为(3.76±0.00) μm和(4.17±0.05) μm(图3 A),平均体积分别为(31.76±0.27) fL和(41.87±1.19) fL。与对照组pH7.10相比,经模拟酸雨处理后,在pH5.65下,469细胞粒径和体积显著降低了2.70%(P<0.05)和11.96%(P<0.05),905则显著降低了3.19%(P<0.05)和11.19%(P<0.05);在pH4.50下,469细胞粒径和体积显著降低了11.39%(P<0.05)和29.93%(P<0.05),905显著降低了3.99%(P<0.05)和14.55%(P<0.05)(图3)。
对比469和905,各pH处理下905藻体细胞粒径和体积显著高于469细胞(P<0.05)。

图3 铜绿微囊藻469与905在pH 7.10以及模拟酸雨(pH5.65和pH4.50)处理下24h后细胞平均粒径大小及平均细胞体积(n=3)Fig.3 The mean cell size and mean cell volume of Mycrocystis aeruginosa strains of 469 and 905 after cultured under pH 7.10and simulated acid rain (pH 5.65 and pH4.50) condition for 24h (n=3)
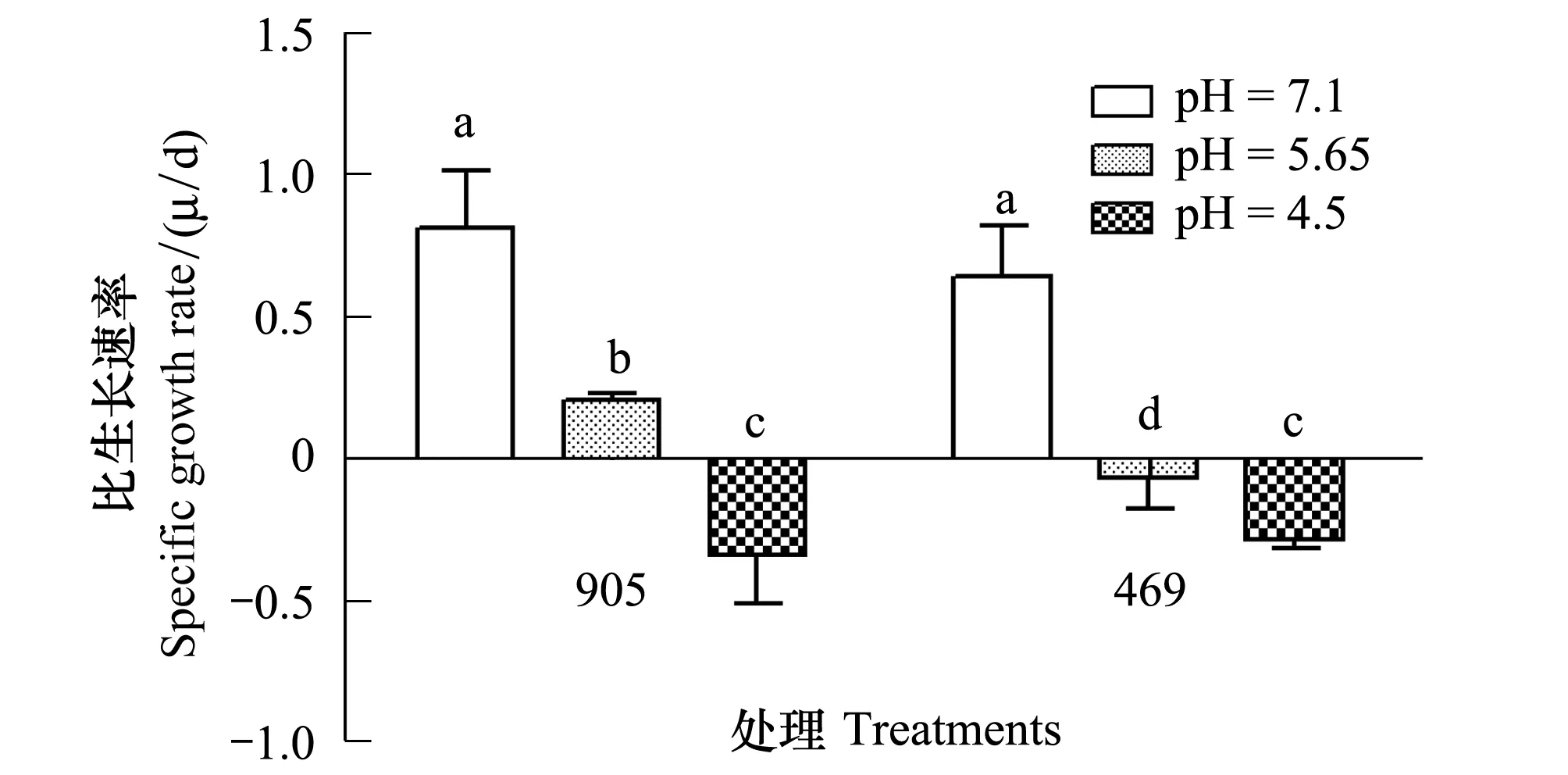
图4 铜绿微囊藻469与905在pH 7.10以及模拟酸雨(pH5.65及pH4.50)处理下24h的比生长速率(n=3)Fig.4 The specific growth rate of Mycrocystis aeruginosa strains of 469 and 905 after cultured under pH 7.10and simulated acid rain (pH5.65 and pH4.50) condition for 24h (n=3)
2.3 生长速率
铜绿微囊藻469和905在经不同pH的模拟酸雨处理后,生长速率都显著降低。相对于pH 7.10处理,pH5.65下生长速率在469和905下分别降低了111.12%(P<0.05)和74.41%,pH 4.50下则分别降低了144.77%和142.17%(图4)。
对比469与905,pH5.65下469细胞生长速率呈负增长,905生长速率显著高于469(3.92倍,P<0.05)(图4)。通过显微镜下对各pH处理的细胞形态进行观察,发现模拟酸雨处理导致469和905藻体细胞破裂或变形。
2.4 有效光化学效率
各pH下培养24h后,相对于905株系,469株系藻体有效光化学效率(Yield)显著高于905株系,在pH为7.10和5.65下分别增加了22.62%(P<0.05)和4.74%(P<0.05);然而,在pH 4.50下藻体Yield显著低于905株系,降低了44.90%(P<0.05)(图5)。
将各pH下培养的藻体置于阳光模拟器下,经PAR及PAB辐射处理1h过程中Yield的变化曲线在两株系间呈不同趋势。905藻体在PAR及PAB下,随培养pH的降低,Yield呈缓慢降低趋势,各pH处理间Yield差异显著(多因素方差分析,P<0.001)。在469株系中,藻体经PAR及PAB辐射处理1h过程中,初始的15分钟辐射处理,Yield在各pH下都呈快速、显著降低趋势(P<0.05);在PAR处理下,较低的pH(4.50)处理,藻体Yield显著低于pH7.10和5.65处理组,而在PAB辐射处理下,各pH下培养的藻体Yield进一步地降低(多因素方差分析,P<0.001)(图5)。
通过对两株系在各pH及两辐射处理下Yield的比值进行计算可得,无论是PAR处理还是PAB处理,905株系藻体Yield都要显著高于469株系(比值大于1),且随pH的降低,比值逐渐升高(图6)。
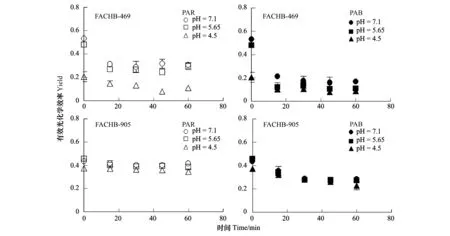
图5 铜绿微囊藻469与905在pH 7.10以及模拟酸雨(pH5.65及pH4.50)处理下24h后置于阳光模拟器下给予可见光(PAR)及可见光+紫外线A+紫外线B(PAB)辐射处理1h过程中有效光化学效率的变化情况(n=3)Fig.5 The Yield value of 469 and 905 strain of Mycrocystis aeruginosa cultured under pH 7.10and simulated acid rain (pH5.65 and pH4.50) condition for 24h when transferred to PAR and PAB for 1 hour exposure (n=3)

图6 铜绿微囊藻469与905在pH 7.10以及模拟酸雨pH5.65及pH4.50下处理24h后,置于阳光模拟器下给予可见光(PAR)及可见光+紫外线A+紫外线B(PAB)辐射处理1h过程中有效光化学效率的比值。虚线表示比值为1(n=3)Fig.6 The Yield ratio of Mycrocystis aeruginosa strains of 469 and 905 cultured under pH of 7.10and simulated acid rain with pH5.65 and pH4.50condition for 24h when transferred to PAR and PAB for 1 hour exposure. The dot line represent the ratio value at 1 (n=3)
图7显示的是pH 7.10、5.65及4.50下两株系经PAR及PAB辐射处理后Yield的抑制率(与各自培养条件下的Yield进行比较)。结果显示低pH(4.50)培养的藻体,无论是905株系还是469株系,辐射处理导致的抑制率都呈升高趋势(图7),且在各pH处理下,PAR(图7)以及PAB(图7)辐射处理1h,905株系的抑制率显著低于469株系(P<0.05)。
3 讨论
微囊藻是我国多数湖泊(如太湖、巢湖、滇池等)发生水华时的优势种[26],包括产毒株和不产毒株,其中产生毒素的微囊藻类群通过释放各种类型的微囊藻毒素,影响水体其他生物代谢活动,并可对人类赖以生存的饮用水源产生潜在影响[27]。以往的原位观测研究表明微囊藻水华发生时多以产毒类型为主[22],究其原因可能与不同类型的藻株对环境变化的适应能力不同导致,产毒与不产毒株系对环境变化的适应能力,可能是决定各微囊藻类群在自然界的分布和丰度的主要因素。因而,在全球气候变化背景下从生理学的角度比较研究不同类型水华蓝藻响应两种环境胁迫(酸雨,UV)的异同,具有较高的研究价值。
本研究结果显示,铜绿微囊藻905与469在模拟酸雨处理下细胞粒径变小,叶绿素含量升高,光化学效率显著降低,并在高光及紫外辐射下表现为进一步地降低;同时两藻株对酸雨及紫外辐射的耐受能力有着显著差异,表现为905藻株更为耐受酸雨胁迫以及紫外辐射。
多数研究表明酸雨可影响浮游植物群落结构[8],然而涉及酸雨对水体典型浮游植物光合生理影响的研究,报导较少。本研究发现,酸雨处理可显著降低细胞粒径和体积,而单位细胞叶绿素含量显著升高。早期研究表明在长期酸性胁迫下,浮游植物叶绿素含量显著受到抑制[28];然而也有研究表明H2SO4型湖泊中,中尺度的酸化可导致叶绿素b、脱镁叶绿素、叶黄素含量在硅藻、甲藻、绿藻中呈增加趋势,但在极度酸化下类胡萝卜素、叶绿素a、脱镁叶绿素则显著降低[29]。通过对细胞生长速率的计算以及显微镜下观察,推测单位细胞叶绿素含量的升高(累积)可能是由于瞬时的酸雨胁迫处理导致细胞分裂速率延缓所引起。通常水体中浮游植物经历恒定的pH变化范围(日变化),酸雨引起的水体H+浓度增加(瞬时酸性胁迫),可影响浮游植物细胞内外的酸碱平衡,例如一些研究发现水体pH降低(H+浓度增加)可提高浮游植物的能量消耗[30],导致细胞呼吸速率在酸性环境下显著升高[31],从而可能使细胞生长速率受到抑制。相比于pH7.10处理,在pH5.65处理下,905藻株生长速率的降低程度低于469藻株,暗示该藻株对水体酸化的耐受性较高。
酸雨发生的同时,可导致水体溶解有机质含量降低,使阳光紫外辐射的透射深度增加,因而酸雨诱导的水体生态系统的改变,与水体紫外辐射的增强有着极大联系[16]。其中紫外波段的光,尤其是UVB部分,可显著抑制浮游植物的光合作用速率[32],降低光合固碳量[33]。紫外辐射增强对水体蓝藻的影响已开展较多研究,内容主要包括UVB辐射对生物大分子的损伤效应[34],对藻体细胞形态[19]、光合色素[35]、生长[20]、固氮[36]、光系统的损伤与修复[21]等方面的效应,然而从酸雨与UV辐射的耦合效应的角度对铜绿微囊藻进行研究,尚未见相关报道。本文的研究表明紫外辐射显著抑制了两株铜绿微囊藻的有效光化学效率,UV辐射导致的光合活性降低在其他蓝藻中也有发现,例如聚球藻经高强度的UVB辐射处理后,藻体PSⅡ活性下降,表现为电子传递速率显著受到抑制[37]。UV辐射导致的光合活性的降低,最终可使蓝藻生长速率受到抑制,这种降低的生长速率在念珠藻、织线藻以及隐杆藻中都有发现[20]。经酸雨胁迫处理后,藻体对紫外辐射的敏感性显著增加(Yield降低,抑制率增加),表明酸雨与UV辐射的耦合可进一步降低浮游植物的光合能力。然而,在响应高光强以及紫外辐射时,905和469藻株光合能力表现出明显的差异(469抑制率更高,更为敏感),表明905藻株更为耐受紫外辐射的胁迫,这一耐受性可能与藻体较高的光保护色素含量有着直接关系[38]。本研究结果也表明,905藻株类胡萝卜素以及紫外吸收物质的含量在各pH处理下都显著高于469藻株,且酸雨胁迫促进(类胡萝卜素)或并未(紫外吸收物质)影响光保护色素的含量。另一方面,光系统反应中心蛋白(D1/D2)在经历高光强或高的UV辐射后,其损伤后修复速率的快慢[39],也是决定二者对UV辐射敏感性不同的另一个可能因素,通过对PSⅡ活性(Yield)的差异进行比较可以看出,905较469可能具有较高的D1/D2蛋白周转速率。
在未来全球变化背景下,随着酸雨的持续影响以及阳光紫外辐射的增强,淡水水体浮游植物将会面临复杂的环境变化,这些环境因子共同作用,对浮游植物的光合生理以及群落结构都将会产生很大的影响,这种影响在很大程度上取决于不同藻体对这些环境变化的响应。本研究中,酸雨及紫外辐射处理对两株藻体的光合生理都产生了较大影响,然而905藻株较强的耐受酸雨以及紫外辐射的能力,可能会使该株系成为淡水水体主要的优势种。未来,结合环境变化研究铜绿微囊藻不同株系之间以与其他浮游植物类群间的关系对预测水体浮游植物群落结构变动具有一定的指导意义。
致谢:感谢温州大学马增岭副教授惠赠藻株、淮海工学院徐军田副教授在论文写作上的帮助。
[1] Felten V, Tixier G, Guérold F. Acid rain ecotoxicity // Férard J F, Blaise C. Encyclopedia of Aquatic Ecotoxicology. The Netherlands:Springer, 2013:1- 14.
[2] Schindler D W. Effects of acid rain on freshwater ecosystems. Science, 1988, 239(4836):149- 157.
[3] Poléo A B S. Aluminium polymerization-a mechanism of acute toxicity of aqueous aluminium to fish. Aquatic Toxicology, 1995, 31(4):347- 356.
[4] Gensemer R W, Playle R C. The bioavailability and toxicity of aluminum in aquatic environments. Critical Reviews in Environmental Science and Technology, 1999, 29(4):315- 450.
[5] Verbost P M, Van Rooij J, Flik G, Lock R A C, Bonga S E W. The movement of cadmium through freshwater trout branchial epithelium and its interference with calcium transport. Journal of Experimental Biology, 1989, 145(1):185- 197.
[6] Jensen F B, Malte H. Acid-base and electrolyte regulation, and haemolymph gas transport in crayfish, Astacus astacus, exposed to soft, acid water with and without aluminium. Journal of Comparative Physiology B, 1990, 160(5):483- 490.
[7] Manny B A, Fahnenstiel G L, Gardner W S. Acid rain stimulation of Lake Michigan phytoplankton growth. Journal of Great Lakes Research, 1987, 13(2):218- 223.
[8] Raut R, Sharma S, Bajracharya R M. Biotic response to acidification of lakes-A review. Kathmandu University Journal of Science, Engineering and Technology, 2012, 8(1):171- 184.
[9] Lessmann D, Fyson R, Nixdorf B. Phytoplankton of the extremely acidic mining lakes of Lusatia (Germany) with pH≤3. Hydrobiologia, 2000, 433(1/3):123- 128.
[10] 姜海波. UV-B 增强对水华蓝藻铜绿微囊藻的长期和短期效应及其光合作用适应 [D]. 武汉:华中师范大学, 2005.
[11] Beardall J, Sobrino C, Stojkovic S. Interactions between the impacts of ultraviolet radiation, elevated CO2, and nutrient limitation on marine primary producers. Photochemical & Photobiological Sciences, 2009, 8(9):1257- 1265.
[12] Torremorell A, Bustigorry J, Escaray R, Zagarese H E. Seasonal dynamics of a large, shallow lake, laguna Chascomús:The role of light limitation and other physical variables. Limnologica-Ecology and Management of Inland Waters, 2007, 37(1):100- 108.
[13] Manney G L, Santee M L, Rex M, Livesey N J, Pitts M C, Veefkind P, Nash E R, Wohltmann I, Lehmann R, Froidevaux L, Poole L R, Schoeberl M R, Haffner D P, Davies J, Dorokhov V, Gernandt H, Johnson B, Kivi R, Kyrö E, Larsen N, Levelt P F, Makshtas A, McElroy C T, Nakajima H, Parrondo M C, Tarasick D W, von der Gathen P, Walker K A, Zinoviev N S. Unprecedented Arctic ozone loss in 2011. Nature, 2011, 478(7370):469- 475.
[14] Zepp R G, Erickson Iii D J, Paul N D, Sulzberger B. Effects of solar UV radiation and climate change on biogeochemical cycling:interactions and feedbacks. Photochemical & Photobiological Sciences, 2011, 10(2):261- 279.
[15] Häder D-P, Helbling E W, Williamson C E, Worrest R C. Effects of UV radiation on aquatic ecosystems and interactions with climate change. Photochemical & Photobiological Sciences, 2011, 10(2):242- 260.
[16] Schindler D W, Curtis P J, Parker B R, Stainton M P. Consequences of climate warming and lake acidification for UV-B penetration in North American boreal lakes. Nature, 1996, 379(6567):705- 708.
[17] Beardall J, Stojkovic S, Gao K S. Interactive effects of nutrient supply and other environmental factors on the sensitivity of marine primary producers to ultraviolet radiation:implications for the impacts of global change. Aquatic Biology, 2014, 22:5- 23.
[18] Whitton B A, Potts M. Introduction to the cyanobacteria // Whitton B A, Potts M. The Ecology of Cyanobacteria:Their Diversity in the Time and Space. Netherlands:Springer, 2000:1- 11.
[19] Ma Z L, Gao K S. Photoregulation of morphological structure and its physiological relevance in the cyanobacteriumArthrospira(Spirulina)platensis. Planta, 2009, 230(2):329- 337.
[20] Zeeshan M, Prasad S. Differential response of growth, photosynthesis, antioxidant enzymes and lipid peroxidation to UV-B radiation in three cyanobacteria. South African Journal of Botany, 2009, 75(3):466- 474.
[21] Jiang H B, Qiu B S. Inhibition of photosynthesis by UV-B exposure and its repair in the bloom-forming cyanobacteriumMicrocystisaeruginosa. Journal of Applied Phycology, 2011, 23(4):691- 696.
[22] 徐瑶. 富营养化水体微囊藻分子生态研究-以太湖和秦淮河为例[D]. 南京:南京师范大学, 2011.
[23] Porra R J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynthesis Research, 2002, 73(1- 3):149- 156.
[24] Parsons T R, Strickland J D H. Discussion of spectrophotometric determination of marine-plant pigments, with revised equations for ascertaining chlorophylls and carotenoids. Journal of Marine Research, 1963, 21(3):155- 163.
[25] 徐军田. 经济红藻龙须菜和海萝的光生物学研究[D]. 汕头:汕头大学, 2008.
[26] 闫海, 潘纲, 张明明. 微囊藻毒素研究进展. 生态学报, 2002, 22(11):1968- 1975.
[27] Codd G A, Bell S G, Brooks W P. Cyanobacterial toxins in water. Water Science & Technology, 1989, 21(3):1- 13.
[28] Kwiatkowski R E, Roff J C. Effects of acidity on the phytoplankton and primary productivity of selected northern Ontario lakes. Canadian Journal of Botany, 1976, 54(22):2546- 2561.
[29] Leavitt P R, Findlay D L, Hall R I, Smol J P. Algal responses to dissolved organic carbon loss and pH decline during whole-lake acidification:Evidence from paleolimnology. Limnology and Oceanography, 1999, 44(3):757- 773.
[30] Wu Y P, Gao K S, Riebesell U. CO2-induced seawater acidification affects physiological performance of the marine diatomPhaeodactylumtricornutum. Biogeosciences, 2010, 7(9):2915- 2923.
[31] Li W, Gao K S, Beardall J. Interactive effects of ocean acidification and nitrogen-limitation on the diatomPhaeodactylumtricornutum. Plos One, 2012, 7(12):e51590.
[32] Guan W C, Gao K S. Impacts of UV radiation on photosynthesis and growth of the coccolithophoreEmilianiahuxleyi(Haptophyceae). Environmental and Experimental Botany, 2010, 67(3):502- 508.
[33] Li G, Gao K S, Gao G. Differential impacts of solar UV radiation on photosynthetic carbon fixation from the coastal to offshore surface waters in the South China Sea. Photochemistry and Photobiology, 2011, 87(2):329- 334.
[34] Sinha R P, Dautz M, Häder D P. A simple and efficient method for the quantitative analysis of thymine dimers in cyanobacteria, phytoplankton and macroalgae. Acta Protozoologica, 2001, 40(3):187- 196.
[35] Ehling-Schulz M, Bilger W, Scherer S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacteriumNostoccommune. Journal of Bacteriology, 1997, 179(6):1940- 1945.
[36] Kumar A, Tyagi M B, Jha P N, Srinivas G, Singh A. Inactivation of cyanobacterial nitrogenase after exposure to ultraviolet-B radiation. Current Microbiology, 2003, 46(5):380- 384.
[37] Prabha G L, Kulandaivelu G. Induced UV-B resistance against photosynthesis damage by adaptive mutagenesis inSynechococcusPCC 7942. Plant Science, 2002, 162(5):663- 669.
[38] Chen S W, Gao K S. Solar ultraviolet radiation and CO2-induced ocean acidification interacts to influence the photosynthetic performance of the red tide algaPhaeocystisglobosa(Prymnesiophyceae). Hydrobiologia, 2011, 675(1):105- 117.
[39] Wu H Y, Abasova L, Cheregi O, Deák Z, Gao K, Vass I. D1 protein turnover is involved in protection of Photosystem II against UV-B induced damage in the cyanobacteriumArthrospira(Spirulina)platensis. Journal of Photochemistry and Photobiology B:Biology, 2011, 104(1- 2):320- 325.
Physiological responses of toxigenic and non-toxigenic strains ofMicrocystisaeruginosato simulated acid rain and UV radiation
LI Wei1,2, YANG Yuling1, HUANG Song1, DONG Lili1, PAN Jian1, LI Yahe2,3, ZHOU Yue1, GAO Kunshan2,*
1CollegeofLifeandEnvironmentalSciences,HuangshanUniversity,Huangshan245041,China2StateKeyLaboratoryofMarineEnvironmentalScience,XiamenUniversity,Xiamen361005,China3CollegeofMarineSciences,NingboUniversity,Ningbo315211,China
Acid precipitation from natural and human activities can lead to different extents of acidification in different freshwater ecosystems, with noticeable changes in the carbonate system and aquatic chemistry. For example, acidification is known to decrease the ionic concentrations of magnesium and calcium, but enhance aluminum concentrations. The changes to chemicals associated with freshwater acidification could induce physiological disturbance to most aquatic organisms, which would then need to spend additional energy to cope with acidic stress. Phytoplankton species with different sensitivities to acid stress may have differential physiological responses; thus, acid stress may lead to altered community structure and species richness. Alternatively, UV-B (280—320nm) irradiance has been increasing due to stratospheric ozone depletion, which would also impact freshwater phytoplankton. Meanwhile, significantly decreased dissolved organic matter as a result of acid rain may further enhance the penetration of solar UV radiation (UVR, 280—400nm) in the water column due to decreased light attenuation. Therefore, freshwater acidification and solar UVR may interact to influence phytoplankton physiology. This study examined the physiological responses of a freshwater bloom forming cyanobacterium,Microcystisaeruginosa, to acidification and UVR. Two strains were used for the experiments; a toxigenic strain (FACHB-905) and a non-toxigenic strain (FACHB-469).Microcystisaeruginosacells were grown under simulated acid rain (with a mole ratio of H2SO4∶HNO3= 8∶1) under a solar simulator with and without UVR, to observe whether photosynthetic performance and sensitivity to UV irradiance differed between the two strains. Three pH levels were used, pH 7.10(regular BG11+N medium), 5.65, and 4.50, with the latter two pH levels reflecting acidification conditions. Two solar radiation treatments were carried out, including:photosynthetically active radiation alone (PAR, 400—700nm) and PAR+ UV-A+UV-B (PAB, 295—700nm). Our results indicate that the mean cell size and cell volume of the 905 strain was significantly higher than those of the 469 strain in all pH treatments. Acidification treatment significantly lowered mean cell size and volume, but enhanced the chlorophyll a content in both strains. Under the lowest pH (4.50), higher mortality rates were observed for both strains, with decreased effective quantumyield. The inhibition of the growth rate was more pronounced in the non-toxigenic strain. The effective quantumyielddeclined with decreasing pH level when cells were exposed to high levels of PAR and PAR+UVR. UV-induced inhibition of effective quantumyieldwas much higher in the non-toxigenic strain compared to the toxigenic strain. This differential response may be attributed to different contents to UV absorbing compounds and carotenoids. Under the global climate change scenario, increasing aquatic acidification and UV irradiance might decrease the competence of the non-toxigenic strain over the toxigenic one.
Microcystisaeruginosa; acid rain; UV radiation; growth; photosynthesis
安徽省自然科学基金青年项目(1508085QC67);黄山学院博士启动项目(2014xkjq002);安徽省大学生创新创业项目(AH2014103753126);近海海洋环境科学国家重点实验室(厦门大学)访问学者基金(MEL Visiting Fellowship) (MELRS1314)
2014- 05- 21; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 19
10.5846/stxb201405211049
*通讯作者Corresponding author.E-mail:ksgao@xmu.edu.cn
李伟,杨雨玲,黄松,董丽丽,潘健,李亚鹤,周月,高坤山.产毒与不产毒铜绿微囊藻对模拟酸雨及紫外辐射的生理响应.生态学报,2015,35(23):7615- 7624.
Li W, Yang Y L, Huang S, Dong L L, Pan J, LI Y H, Zhou Y, Gao K S.Physiological responses of toxigenic and non-toxigenic strains ofMicrocystisaeruginosato simulated acid rain and UV radiation.Acta Ecologica Sinica,2015,35(23):7615- 7624.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
浙江气象(2019年1期)2019-04-02
科技创新与应用(2017年20期)2017-07-15
环境保护与循环经济(2017年8期)2017-03-22
装备环境工程(2015年4期)2015-02-28
西北园艺(果树)(2015年1期)2015-02-21
湖泊科学(2015年4期)2015-02-17
应用海洋学学报(2014年2期)2014-11-26
癌变·畸变·突变(2014年2期)2014-03-01
华东理工大学学报(自然科学版)(2014年3期)2014-02-27
