
马铃薯ACS基因克隆及生物信息学分析
2015-01-18 05:46裴瑞芳司怀军
甘肃农业大学学报 2015年1期
刘 英,裴瑞芳,王 洁,张 宁,司怀军,王 蒂
(1.甘肃省作物遗传改良与种质创新重点实验室,甘肃省干旱生境作物学省部共建国家重点实验室培育基地,甘肃 兰州 730070;2.甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
乙烯作为一种植物气体激素,对植物种子萌发、生长发育、植物衰老、果实成熟等有明显的影响,并参与了从种子萌发到成熟衰老一系列生命过程的调节.同时与植物逆境胁迫如机械伤害、冷害、渍水、病原入侵等方面有关[1,2],在接种晚疫病后 ACS表达量显著升高[3];此外,乙烯还参与植物细胞的程序性死亡,促进花凋谢和DNA降解.有报道称乙烯与马铃薯块茎萌发有密切关系[4],控制乙烯的生物合成就可以提前或推迟发芽时期,从而来延长或缩短马铃薯块茎休眠时间.
在高等植物体内,乙烯合成需要2个关键酶[5]:ACC合成酶(ACC synthase,ACS)和 ACC氧化酶(ACC oxidase,ACO).ACC合成酶是整个乙烯合成途径中的限速酶,催化S-腺苷甲硫氨酸(SAM)产生1-氨基环丙烷-1-羧酸(1-aminocyclo-propane-1-carboxylic acid,ACC),ACC在 ACC氧化酶作用下产生乙烯[6].ACC合成酶(ACS)是以磷酸吡哆醛为辅酶,可能存在于植物胞质中,存在形式有单体、二聚体和三聚体3种形式,其中单体ACS活性较高,在反应中不易失活[7].受多种因素调控,在植物组织中含量很低,因此分离纯化比较困难.
研究表明,ACS有许多基因编码,其中在番茄(Lycopersicon esculentum)和拟南芥(Arabidopsis thaliana)中分别分离到9个和12个,而苹果(Malusxdomestica)中也至少存在4个该基因成员[8-9].目前已经从番茄、苹果、康乃馨、豇豆、笋瓜、冬瓜、桃、猕猴桃、香蕉、柑橘、哈密瓜、柿、玫瑰、菠萝、枇杷还有拟南芥中克隆出ACS基因[10].
本研究克隆了马铃薯ACS基因cDNA全长序列,并利用生物信息学软件分析ACS基因结构,而且预测了ACS基因编码蛋白的物理性质,为下一步利用基因工程技术,通过正义过表达和RNAi调控ACS合成,进而调控乙烯对马铃薯块茎休眠的生理效应,延长或缩短马铃薯块茎休眠时间奠定了一定基础.
1 材料和方法
1.1 材料与试剂
马铃薯四倍体栽培品种‘甘农薯2号’试管苗、大肠杆菌(Escherichia coli)DH5α由甘肃省作物遗传改良与种质创新重点实验室保存.PMD18-T载体、T4连接酶和 LA Taq DNA 聚合酶、SacⅠ、BamH Ⅰ、XhoⅠ、XbaⅠ等限制性内切酶均购于TaKaRa公 司;RNA simple Total RNA kit购 自TIANGEN公司;M-MLV逆转录酶、胶回收试剂盒购自上海生物工程技术服务有限公司,引物也由该公司合成.其他试剂均为国产或进口分析纯试剂.
1.2 试验方法
1.2.1 马铃薯叶片总RNA提取与cDNA合成利用植物总RNA提取试剂盒提取马铃薯试管薯叶片总RNA,各步骤均按试剂盒说明书进行.用琼脂糖凝胶电泳检测提取RNA质量,并选用高质量的RNA进行下游试验.利用M-MLV逆转录酶合成cDNA第1链,具体操作过程参照试剂盒说明书.cDNA保存于-20℃,用于后续试验.
1.2.2 马铃薯ACS基因引物设计及克隆 通过已报道的马铃薯 ACS 基因(序列号为PGSC0003DMT400014932)的序列,利用 Primer 5.0设计PCR引物Ps(上游引物F:GGGATCC(BamH Ⅰ)AAATGGGGTTAATTTCAG;下游引物R:CGAGCTC(SacⅠ)GAACATCACGATTAATTC),由上海生物工程技术服务有限公司合成.其中ACS上下游引物分别添加BamH Ⅰ、SacⅠ酶切位点.
PCR反应体系为25μL,10×LA Taq Buffer 2.5μL,dNTP Mixture(2.5mmol/L)4μl,上、下游引物各1μl,模板cDNA 2μL,LA Taq聚合酶0.25μL,ddH2O 14.25μL.反应条件为:94℃预变性3min;94 ℃变性45s,58℃退火1min 30s;72℃延伸1min 30s,35个循环;72℃延伸10min.目的产物用琼脂糖凝胶电泳检测,经回收纯化,克隆到PMD18-T载体上,选择阳性克隆,经PCR、酶切验证正确后送上海生物工程技术服务有限公司测序.
1.2.3 基因生物信息学分析 依据ACS核苷酸和氨基酸序列,利用DNAMAN,DNAstar等生物信息学软件进行分析.利用NCBI和DNAMAN完成核算及氨基酸序列的组成成分、相对分子质量及等电点理化性质分析;采用NCBI在线工具完成核算及氨基酸序列的同源性比对、多序列比对,使用MEGA 4软件进行系统进化树构建;DNAstar和THMM工具完成亲水性/疏水性、跨膜结构域的在线分析;通过蛋白质专家系统的在线工具(http://www.expasy.org/)分别完成对蛋白质二级、三级结构的预测.
2 结果与分析
2.1 ACS基因的克隆
本研究用TIANGEN公司的RNA simple To-tal RNA kit总RNA提取试剂盒提取马铃薯叶片总RNA,经琼脂糖凝胶电泳检测总 RNA 3条带(28S、18S、5S)条带清晰、完整.以cDNA 为模板,PCR扩增结果如下.ACS PCR扩增产物为1 474bp(图1),与预期大小相同.
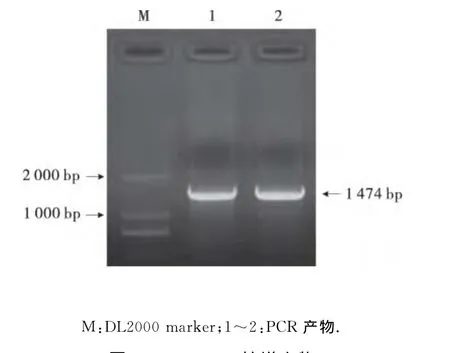
图1 ACS PCR扩增产物Fig.1 Amplified PCR products of ACSgene
2.2 马铃薯ACS基因的生物信息学分析
2.2.1 铃薯ACS基因序列分析 ACS mRNA全长1 832bp其中CDS区长1 461bp.通过 NCBI的blast比对发现,马铃薯ACS基因与茄科中的番茄(Lycopersicon esculentum,GI:33411116)同 源 性 达97%,与烟草(Nicotiana glutinosa,GI:3108180)ACS基因同源性达98%,与辣椒(Capsicum chinense,GI:l 186200769)ACS基因比对同源性达84%.
2.2.2 马铃薯ACS蛋白序列分析 ACS共有486氨基酸残基,分子量约为55kU,PI 6.76.对ACS的氨基酸组成进行分析,发现该蛋白质中色氨酸含量最少为6个,亮氨酸含量最高为41个.
2.2.3 不同植物ACS蛋白质序列比对 利用NCBI上的blast程序分别选取高等植物和低等植物中的ACS的同源蛋白,序列比对结果如图2.分析结果表明,ACS在高等植物中保守性较高,可达到92%.

图2 马铃薯ACS蛋白同源比对Fig.2 Comparison of the amino acid sequences of ACS
2.2.4 同其他物种的进化关系聚类分析 在NCBI中查找其他物种ACS基因编码的氨基酸序列,并利用 MEGA 4软件将马铃薯(Solanum tuberosum)ACS基因编码的氨基酸序列与茄科中番茄(Solanum lycopersicum,GI:3986117)、烟草(Nicotiana sylvestris,GI:3108181)、辣椒(Capsicum annuum,GI:186200770)、毛果杨(Populus trichocarpa,GI:224053855)、葡 萄 (Vitis vinifera,GI:359474171)、水稻(Oryza sativa,GI:115460074)、拟南芥(Arabidopsis thaliana,GI:15237126)、大豆(Glycine max,GI:351727557)、苔 藓 (Physcomitrella patens,GI:168051669)、衣 藻 (Chlamydomonas reinhardtii,GI:159466366)进行系统进化分析,构建了系统进化树.结果显示马铃薯和茄科中番茄同源蛋白在进化上属于同一个分支,分子进化最近,关系最为密切(图3).

图3 马铃薯ACS蛋白的进化树分析Fig.3 The potato ACS protein phylogenetic analysis
2.2.5 马铃薯ACS其他二级结构亲水性和疏水性分析 对ACS的二级结构域、疏水性和亲水性进行分析,结果如图4.ACS具有12个alpha螺旋区域,22个beta片层区,32个转角和23个自由卷曲区.具有39个疏水区和41个亲水区.
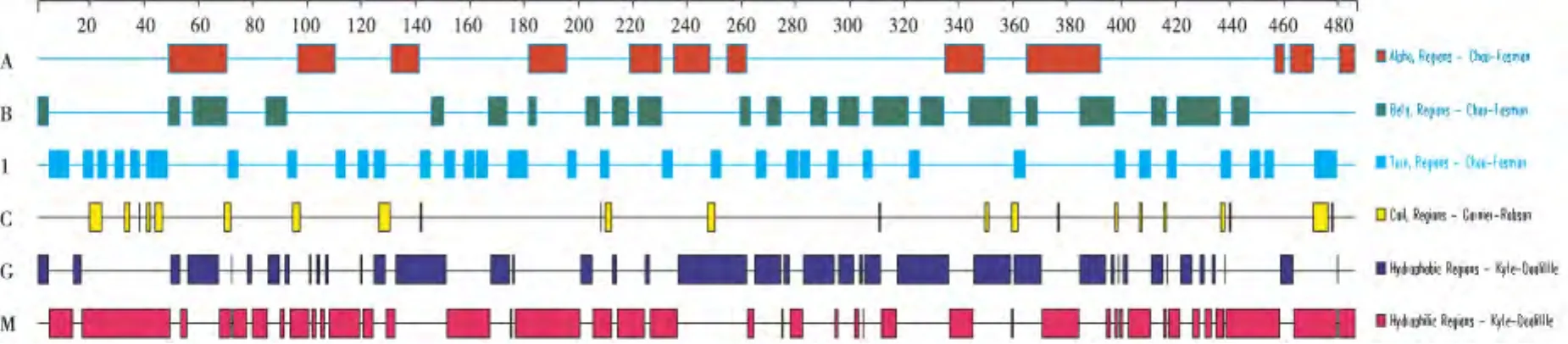
图4 马铃薯ACS蛋白二级结构、亲水性和疏水性分析Fig.4 Analysis of secondary structure,hydrophile and hydrophobicity of potato ACS
2.2.6 马铃薯ACS蛋白质三级结构预测 对ACS的三级结构进行预测(图5).返回的有效结果显示,ACS是以双体形式存在的.统计三维结构中alpha螺旋和beta片层的数量发现,和二级结构预测的结果不能完全匹配.通过3D结构分析,ACS的反应中心可能是在二聚体组成的中心空隙中.
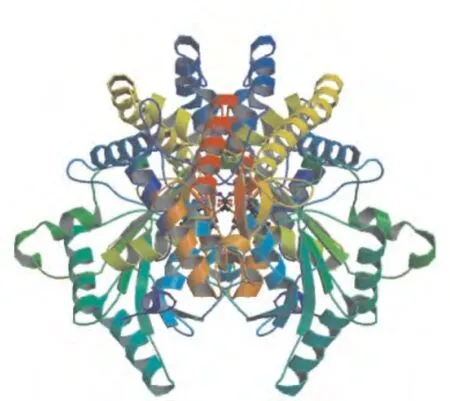
图5 马铃薯ACS蛋白三级结构预测Fig.5 The tertiary structure prediction of potato ACS
3 讨论
马铃薯块茎的储存在生产加工过程中面临很大挑战.即使处于适宜的外界环境中,成熟的马铃薯块茎不会发芽.块茎顶芽的这种休眠(又称为块茎休眠)利弊兼存:有利的方面是,植物在不适宜的环境中仍可以生存,而且对人们而言收获块茎后需要长期储存时也是非常有益的;但是收获后需要尽快播种时,又是非常不利的.气候环境不允许马铃薯的全年种植,长期储存也是不可避免的.通常储存方法有2种:低温储存或使用萌发抑制剂如间氯苯氨甲酸异丙酯(CIPC)及马来酰肼处理来延迟发芽.这2种方法也都存在弊端:抑制剂处理时要非常小心,因为萌发抑制剂同时也是细胞分裂的强抑制剂;马来酰肼使用不当也会限制马铃薯块茎大小和产量;CIPC处理后会有长达一年之久的残留,需要对残留物进行彻底清除.此外,抑制剂的处理可能回对环境和人体健康产生一定的影响.低温储存相对容易操作,但经常会导致块茎中淀粉降解,不仅使块茎产生甜味,而且工业加工过程中可能产生“美拉德反应”.因此,通过转基因方法,开辟一条有效的途径,来延长或缩短马铃薯块茎休眠时间,为种植和加工行业提供工具,也希望通过本研究能对马铃薯新品种培育方面做出一定贡献.
马铃薯休眠与乙烯合成密切相关.近年来,对乙烯合成过程中ACS做了深入的研究,并取得了一些突破性的进展.1986年初次从番茄中纯化得到了ACS,1989年从南瓜中分离出ACS编码基因(Sato等).1990年从番茄cDNA文库中分离出ACS基因序列[11],之后从其它植物中克隆出ACS基因.本试验对乙烯合成过程中的ACS基因进行克隆和生物信息学分析,结果表明,ACS以双体的形式存在[12],在高等植物中保守性较高.在进化上马铃薯ACS和番茄同源蛋白属于同一个分支,分子进化最近,关系最为密切.对马铃薯ACS基因的研究不仅有助于了解乙烯的合成机制,而且为研究该基因在马铃薯生长发育和逆境响应过程中的作用奠定了基础[13-14].试验下一步将构建马铃薯ACS基因植物正义和RNAi表达载体[15-16],并通过农杆菌介导的遗传转化法获得转基因植株,通过正义表达、RNAi等调控乙烯合成,进而调控乙烯对马铃薯植株的生理效应,延长或缩短马铃薯块茎休眠期的程度[17-18].
[1] Bleecker A B,Kende H.Ethylene:agaseous signal molecule in plants[J].Annu Rev Cell Dev Biol,2000,16:1-18
[2] 赵晋锋,余爱丽,王高鸿,等.植物CBL/CIPK网络系统逆境应答研究进展[J].中国农业科技导报,2011,13(4):32-38
[3] 朱佳莉,丁艳,徐宏章,等.接种晚疫病病原菌对马铃薯水杨酸、乙烯合成关建酶基因表达的影响[J].湖北农业科学,2013,52(17):4239-4265
[4] Irena P,Lawrence R,Harlan K P.Dual effects of ethylene on potato dormancy and sprout growth[J].Plant Physiol,1974,53:658-662
[5] Yang S F,Hoffman N E.Ethylene biosynthesis and its regulation in higher plants[J].Ann Rev Plant Physiol,1980,35:155-189
[6] Yoon S K,Hyun S L,Yong H K,et al.Elevated H2O2production via overexpression of a chloroplastic Cu/Zn SOD gene of lily(Lilium oriental hybrid ‘Marco Polo’)triggers ethylene synthesis in transgenic potato[J].Plant Cell,2008,27:973-983
[7] Satoh S.Monomeric and dimeric forms and the mechanismbased inactivation of 1-aminocycloprame-1-carboxylate sythase[J].Plant Cell Physiol,1993,34:753-760
[8] Jakubowicz M,Sadowski J.1-Aminocyclopropane-1-carboxylate synthase genes and expression[J].Acta Physiol,2002,24(4):459-478
[9] Varanasi V,Shin S,Mattheis J,et al.Expression profiles of the MdACS3gene suggest a function as an accelerator of apple(Malus xdomestica)fruit ripening[J].Postharvest Bio Technol,2011,62(2):141-148
[10] 音建华,赖钟雄,林玉玲,等.枇杷胚性培养物中2个ACS基因的克隆及生物信息学分析[J].热带作物学报,2012,33(12):2214-2219
[11] Straeten D V,Wiemeersch L V,Goodman H M,et al.Cloning and sequence of two different cDNAs encoding 1-aminocyclopropane-1-carboxylate synthase in tomato[J].PNAS,1990(87):4859-4863
[12] 王翊,胡宗利,杨妤欣,等.水稻SBP基因家族的生物信息学分析[J].生物信息学,2011,9(1):82-88
[13] Pouvreau B,Baud S,Vcrnoud V,et al.Duplicate maize Wrinkled1transcription factors activate target genes involved in seed oil biosynthesis[J].Plant Physiol,2011,156(2):674-686
[14] Zhang W,Wan X,Hong Y,et al.Plant phospholipase D[M]//Munnik D T.Lipid signaling in plants(plant cell monographs).Berlin:Springer publishing company,2010:39-62
[15] 李竞芸,张广辉,王森.RNA干涉及其在植物改良上的应用[J].分子植物育种,2007,5(1):145-148
[16] Sayaka H,Shin-ichiro O,Eri A,et al.The effects of spacer sequences on silencing efficiency of plant RNAi vectors[J].Plant Cell Reports,2007,26(5):651-659
[17] Ada Linkies,Kerstin Muller,Karl Morris,et al.Ethylene interacts with abscisic acid to regulate endosperm rupture during germination:a comparative approach using Lepidium sativum and Arabidopsis thaliana[J].The Plant Cell,2009,21:3803-3822
[18] Wang M B,Wesley S V,Finnegan E J,et al.Replicating satellite RNA induces sequence-specific DNA methylation and truncated transcripts in plants[J].RNA,2001,7,16-28
猜你喜欢
今日农业(2021年15期)2021-11-26
现代畜牧科技(2021年4期)2021-07-21
中国博物馆(2018年2期)2018-12-05
石油化工建设(2018年2期)2018-07-11
现代园艺(2017年19期)2018-01-19
云南中医学院学报(2015年2期)2015-07-31
现代检验医学杂志(2015年4期)2015-02-06
武夷学院学报(2014年5期)2014-07-19
自动化博览(2014年9期)2014-02-28
无机化学学报(2014年9期)2014-02-28
